
En la biología en general, el término raza se usa para definir grupos de características hereditarias comunes en los que se subdividen algunas especies animales.
Desde la década de 1940, los científicos evolucionistas rechazaron la idea de raza, según la cual un número finito de características esenciales pueden usarse para determinar el número de grupos humanos o tipos humanos. Muchos científicos evolucionistas y sociales opinan que las definiciones de raza relativa a los humanos son imprecisas y arbitrarias.[cita requerida]. Algunos de los antropólogos argumentan que la especie humana está compuesta por una única raza dividida en diferentes etnias que, a su vez, se dividen en pueblos. Estas teorías antropológicas aparecieron en los años 1960, en parte como reacción frente a las teorías pseudocientíficas relativas al predeterminismo biológico que predeterminaron el segregacionismo y la discriminación racial, presentes por aquel tiempo en los países occidentales. También influyeron los resultados de las investigaciones de los antropólogos Franz Boas y Claude Lévi-Strauss, que resaltaron las tendencias etnocéntricas de toda cultura.
En su declaración La cuestión racial, de 1950, la UNESCO recomendó sustituir la noción de raza humana, considerada no científica y confusa, por la de etnia, basada más en las diferencias culturales (lengua, religión, costumbres y otras).
En la antropología moderna, existe un amplio consenso científico de que no existen razas humanas en un sentido biológico.
Las razas no existen, ni biológicamente ni científicamente. Los hombres por su origen común, pertenecen al mismo repertorio genético. Las variaciones que podemos constatar no son el resultado de genes diferentes. Si de "razas" se tratara, hay una sola "raza": la humana.José Marín Gonzáles
Encuesta sobre desacuerdo entre disciplinas
En un debate realizado en la primera década del siglo XXI, una parte de los genetistas exponen que la raza no es un concepto significativo ni un mecanismo heurístico útil, dado que se basa en descripciones físicas aparentes sin fundamento biológico. Se ha concluido que el intento de definir razas implica de alguna manera la existencia de una pureza racial, ilustrada por individuos “tipo”. Los avances de la genética apuntan a que las diferencias genéticas entre grupos son biológicamente insignificantes, en el sentido de que existen más variaciones genéticas al interior de los grupos denominados razas que entre ellos, y que los rasgos raciales se solapan sin fronteras diferenciadas.
En contraste, algunos genetistas sostienen que tanto las categorías de identidad propia de raza como de origen étnico son válidas y útiles, que estas categorías corresponden a ciertos grupos, y que esta correspondencia implica que los factores genéticos pueden contribuir a las variaciones fenotípicas inexplicables entre grupos.
La encuesta más reciente, hecha en 1985 (Lieberman et al. 1992), preguntó a 1200 científicos cuántos no estaban de acuerdo con la siguiente proposición: “Hay razas biológicas en la especie Homo sapiens”. Las respuestas fueron como sigue:
- biólogos 16%
- psicólogos evolutivos 36%
- antropólogos físicos 41%
- antropólogos culturales 53%
Estos datos muestran que una mayoría de ellos sí respaldaban el concepto de razas humanas biológicas. La cifra para los antropólogos físicos en los departamentos donde se otorgan los doctorados fueron ligeramente superiores, donde aumentaron de 41% a 42%, y donde el 50% estaban de acuerdo.[cita requerida]
Historia del concepto de raza
Dadas las complejas relaciones sociales, se supone que los seres humanos siempre se han observado a sí mismos y han especulado sobre las diferencias físicas entre los individuos y los grupos. Pero diferentes sociedades han atribuido diferentes significados a dichas diferencias.
Edad Antigua
La división de la humanidad en distintas «razas» —aunque el término no aparecerá hasta el siglo XV d. C.— se encuentra en el Libro de las puertas, texto sagrado del Antiguo Egipto. Este escrito identifica cuatro categorías convencionalmente etiquetadas como “egipcios”, “asiáticos”, “libios”, y “nubios”. En la base de esta clasificación se mezclaban características como el color de piel con las identidades tribales o «nacionales».
En el mundo greco-romano antiguo el determinismo medioambiental fue ampliamente admitido. Fue expuesto por primera vez en detalle en el tratado atribuido a Hipócrates Aires, aguas, lugares del siglo V a. C.. Así, los asiáticos, que habitan un clima cálido y meridional, serían indolentes y pacíficos, pero inteligentes, mientras que los europeos, que viven en un clima frío y septentrional, serían valientes y belicosos, pero faltos de inteligencia. Este modelo fue desarrollado por Aristóteles en el siglo siguiente en su Política, yendo más allá que Hipócrates en el determinismo medioambiental al señalar que el medio ideal en el que vivían los griegos les predisponía a gobernar a los pueblos menos favorecidos por la naturaleza y que al combinar las mejores cualidades de los europeos y de los asiáticos estaban capacitados para gobernar a la humanidad entera. Sin embargo, hay autores de la Antigüedad clásica como Estrabón, que no tienen en cuenta la teoría medioambiental en su descripción de los pueblos y de sus costumbres.
Edad Media

El cristianismo aportó un nuevo concepto, el universalismo, hasta entonces ajeno a la Antigüedad al considerarse la verdadera religión de toda la humanidad. De esta forma la división entre griegos/romanos y «bárbaros», propia de la Antigüedad, fue sustituida por la diferenciación entre los que ya formaban parte de la comunidad cristiana, los bautizados, y «los todavía no cristianos» (los paganos). Un grupo especial lo constituían los judíos ya que eran la cuna de la religión cristiana y por tanto no eran perseguidos, pero solo «conllevados» que no tolerados. Las regiones desconocidas de la Tierra en el imaginario del Occidente medieval aparecían pobladas por seres fabulosos no destinados a la salvación.
Por otro lado, en el Islam medieval la maldición de Cam, convenientemente reelaborada, fue utilizada para justificar la esclavitud de los negros al señalarlos como los descendientes de Cam que, según el relato bíblico, se había mofado de su padre Noé cuando lo encontró borracho y desnudo y Noé furioso había maldecido al hijo de Cam, Canaan, a ser «para sus hermanos el esclavo de los esclavos». En la Biblia nada se decía del color de la piel de Cam (en realidad se trataba de justificar la esclavitud de los cananeos, los grandes enemigos de Israel), pero en el siglo III el padre de la Iglesia Orígenes añadió a la maldición el prejuicio de la piel al afirmar que los hijos de Cam estaban abocados a una vida degradante marcada por la oscuridad (en sentido espiritual). Será el gran erudito árabe Al-Tabari el que en el siglo X afirmó claramente que la maldición de Cam había acarreado el ennegrecimiento de su piel por lo que sus descendientes eran los negros que estaban condenados a la esclavitud.
Siglos XV, XVI y XVII: el nacimiento del término «raza»
La primera constatación escrita de la palabra «raza» en castellano se encuentra en la obra El Corbacho del arcipreste Alfonso Martínez de Toledo publicada en 1438.
[...] toma dos fijos, uno de un labrador, otro de un cavallero: críense en una montaña so mando e disciplina de un marido e muger. Verás cómo el fijo del labrador todavía se agradará de cosas de aldea, como arar, cavar e traher leña con bestias; e el fijo del cavallero non se cura salvo de andar corriendo a cavallo e traer armas e dar cuchilladas e andar arreado. Esto procura naturaleza; asy lo verás de cada día en los logares do byvieres, que el bueno e de buena rraza todavía rretrae do viene, e el desaventurado, de vil rraza e linaje, por grande que sea e mucho que tenga, nunca rretraerá synón a la vileza donde desciende.
En esta obra el arcipreste de Talavera usa la palabra «raza» como sinónimo de linaje para señalar que un «labrador» será siempre un «labrador» («de vil raza e linaje») y un «cavallero» un «cavallero» («de buena raza») sin importar el contexto social en que se eduquen. Según Max Sebastián Hering Torres, de la Universidad Nacional de Colombia, «en este pasaje se evidencia que el término “raza” no pretendía nada más que ser una manifestación de procedencia, es decir, de linaje. El autor utiliza en principio la expresión “raza” de manera neutral y solo mediante la inclusión de un adjetivo positivo “buena raza” o de uno de carácter negativo “vil raza”, el término obtiene un componente valorativo. La palabra “raza” en sí misma no contiene, por tanto, ni una connotación halagadora ni peyorativa. Igualmente se muestra que dicha concepción de “raza” está acompañada por el imaginario de un ethos natural inmanente e invariable del ser».
Sin embargo, el significado más extendido del término «raza» en castellano en los siglos XV, XVI y XVII no era el de sinónimo de linaje, sino el de «mancha» o linaje «defectuoso», «maculado», o de «sangre impura». Proviene de la aplicación del término «raza» en otros contextos, como lo recoge el humanista Antonio Nebrija en su Diccionario (1495) cuando traduce la locución latina panni raritas como «raça del paño», es decir, una rareza o defecto en los paños, expresión muy utilizada por el gremio de sastres.
Así, los que tenían un linaje sin «impurezas» no tenían «raza». Agustín Salucio en 1599 redactó un discurso contrario a los estatutos de limpieza de sangre en el que en uno de sus apartados decía: «[...] porque para tener raza basta un rebisabuelo judio, aunque los otros 15 sean Cristianisimos y nobilissimos». En 1611 el filólogo Sebastián de Covarrubias en su renombrada obra Tesoro de la lengua castellana o española definía «raza» como «la casta de cauallos castizos, a los quales señalan con hierro para q sean conocidos» y decía también que «raza en los linages se toman en mala parte, como tener alguna raza de Moro, o Judio». Lorenzo Franciosini, posiblemente inspirado en Covarrubias, desarrolló en su libro Vocabolario español, e italiano una definición que pone de manifiesto la cercanía entre «limpieza» y «raza» de la siguiente manera: «Limpio: es a veces utilizado en España. Todo el que es cristiano viejo, es porque no tiene raza, ni procedencia mora ni judía». En 1638 Bartolomé Jiménez Patón escribió: «[...] que son los limpios Christianos viejos, sin raza, macula, ni descendencia, ni fama, ni rumor dello». Max Sebastián Hering Torres concluye que «no existe un nexo semántico-ideológico entre el término “raza” utilizado en los siglos XVI-XVII, con el utilizado en los siglos XVIII-XX».
En inglés no fue sino hasta el siglo XVI cuando se empezó a usar el término race (raza), que proviene del francés race. Es probable que tanto race como raza sean un préstamo del italiano -razza-. Los significados del término en el siglo XVI incluían los “vinos con un sabor característico”, “gente con una ocupación común” y “generación”. No existe certeza sobre el origen prístino de la palabra, aunque se sugiere que pudiera, por ejemplo, ser derivado de la palabra árabe “rā's” (رأس), es decir, “cabeza”, pero también “inicio” u “origen”.
Según algunos autores, la palabra «raza», junto con varias de las ideas asociadas en nuestros días al término, son producto de la era colonialista europea. A medida que los europeos se hallaron frente a frente con personas de diferentes partes del mundo, comenzaron las especulaciones sobre las diferencias físicas, sociales y culturales entre los grupos humanos. Muchas de esas reflexiones quedaron registradas en diarios de viaje o bitácoras de navegación. Al aumentar la trata de esclavos africanos, que vino a incorporarse a un mercado esclavista preexistente, hubo un incentivo más para categorizar los grupos humanos y justificar el trato bárbaro de los esclavos llevados a América. Los europeos dieron en clasificar a los pueblos con los que entraron en contacto según la apariencia física, el comportamiento y las capacidades físicas. Un conjunto de creencias del folclore mantuvieron unidas las diferencias físicas heredadas entre grupos con cualidades intelectuales, de comportamiento y morales Ideas similares pueden encontrarse en otras culturas.
La primera clasificación «cintífica» de los humanos en distintas «razas» podría ser la Nouvelle division de la terre par les différents espèces ou races qui l'habitent (“Nueva división de la Tierra por las diferentes especies o razas que la habitan”), de François Bernier, publicada en 1684.[cita requerida]
Siglo XVIII
En el siglo XVIII, las diferencias entre grupos humanos se convirtieron en el centro de una importante rama de la investigación científica (Todorov 1993). Inicialmente, los eruditos se concentraron en catalogar y describir las variedades naturales de la humanidad. The Natural Varieties of Mankind es el título que Johann Friedrich Blumenbach dio a un texto suyo, escrito en 1775. En él, Blumenbach estableció cinco divisiones amplias de los humanos, mismas que se reflejan en ciertas clasificaciones raciales de las últimas décadas. Durante los siglos XVII al XIX, las creencias populares sobre las diferencias de grupo mezcladas con explicaciones científicas de estas diferencias produjeron lo que se ha llamado ideología de raza (Smedley, 1999). Según este planteamiento, las razas son primordiales, naturales, duraderas y distintas. Algunos grupos pueden ser resultado de la mezcla entre antiguas poblaciones distintas, pero un estudio meticuloso puede distinguir aquellas razas ancestrales que se han combinado para producir grupos mestizos.
Siglo XIX
La comunidad científica del siglo XIX fue testigo de los intentos por cambiar el sentido de la palabra raza, de un concepto taxonómico a uno biológico . En el siglo XIX algunos científicos naturales escribieron sobre el tema, entre ellos Georges Cuvier, Charles Darwin, Alfred Wallace, Francis Galton, James Cowles Pritchard, Louis Agassiz, Charles Pickering y Johann Friedrich Blumenbach. A medida que la antropología fue consolidádose durante el siglo XIX, los científicos europeos y estadounidenses buscaron cada vez más explicaciones para las diferencias culturales y de comportamiento que atribuían a la naturaleza de los grupos (Stanton 1960).
Estos científicos hicieron tres afirmaciones sobre la raza:
- ...primero, que las razas son divisiones objetivas y naturales de la humanidad;
- ...segundo, que hay una fuerte relación entre las razas biológicas y otros fenómenos humanos —como formas de actividad y relaciones interpersonales y cultura, y por extensión el relativo éxito material de las culturas—, de suerte que biologizaron la noción de “raza”, como también haría Michel Foucault en el siglo XX;
- ...y tercero, que la raza es por tanto una categoría científicamente válida que se puede usar para explicar y predecir comportamientos individuales y de grupo. Las razas se distinguieron por el color de piel, tipo de rostro, perfil y tamaño craneal, textura y color del pelo. Además, las razas fueron casi consideradas universalmente reflejo de las diferencias del grupo en el carácter moral e inteligencia.
El movimiento eugenésico de finales del siglo XIX y principios del siglo XX, inspirado por Ensayo sobre la desigualdad de las razas humanas, de Joseph Arthur de Gobineau, o la “antroposociología” de Vacher de Lapouge y las teorías de Johann Gottfried von Herder, sostuvieron la "inferioridad biológica" de algunos grupos particulares (Kevles, 1985). En muchas partes del mundo, la idea de raza se convirtió en una manera de dividir rígidamente grupos por cultura. Pero sobre todo, las clasificaciones raciales se apoyaron en las diferencias físicas evidentes (Hannaford 1996). Las campañas de opresión a menudo recurrían al discurso científico que sostenía la supuesta inferioridad inherente por herencia genética a ciertos grupos para legitimar actos inhumanos contra otros, como el genocidio (Horowitz, 2001) o el etnicidio.
El desarrollo de modelos evolucionistas darwinianos (véase Charles Darwin) y la genética mendeliana (véase Gregor Mendel) llamaron la atención de la validez científica de ambas características, y fueron la base de la reconsideración radical del concepto de "raza".[cita requerida]
Siglo XX
En 1976, Richard Dawkins publicó El gen egoísta, en el cual postulaba su teoría evolutiva enfocada en los genes, con los conceptos de meme y memética. En 1982, en su libro El fenotipo extendido, afirmó que los efectos fenotípicos no están limitados al cuerpo de un organismo, sino que pueden extenderse en el ambiente, incluyendo los cuerpos de otros organismos.[cita requerida]
Debates modernos
Raza como subespecie
Con la llegada del neodarwinismo a principios del siglo XX, los biólogos desarrollaron un nuevo y más riguroso modelo de raza como subespecie. Para estos biólogos, una raza es un grupo reconocible que forma parte de una especie. Una especie monotípica no tiene razas, o más bien una raza contiene a toda la especie. Las especies monotípicas pueden caracterizarse de diferentes maneras:
- Todos los miembros de la especie son tan similares que no pueden dividirse sensiblemente en subcategorías biológicamente significativas.
- Los individuos varían considerablemente pero la variación es esencialmente aleatoria y poco significativa por lo que hace a la transmisión genética de estas variaciones. Muchas especies de plantas encajan en esta categoría, razón de que los horticulturistas, interesados en preservar un color de flor particular, eviten la propagación desde la semilla y en cambio usen métodos como la propagación por cortes.
- La variación entre los individuos es importante y sigue un patrón, pero no hay líneas divisorias claras entre grupos separados: se confunden imperceptiblemente unos con otros. Estas variaciones clinales siempre indican flujos genéticos sustanciales entre grupos aparentemente separados que constituyen una población. Las poblaciones que tienen un flujo genético sostenido y sustancial entre sí son más propensas a presentar especies monotípicas, incluso si la variación genética es obvia a la vista.
Una especie politípica tiene dos o más razas (o, en lenguaje corriente, dos o más subtipos). Esta clasificación refleja grupos separados que son claramente distintos uno del otro y generalmente no se entrecruzan (aunque puede haber una relativa estrecha zona de hibridación), pero que podrían entrecruzarse libremente si se diera el caso. Aunque especies diferentes pueden a veces entrecruzarse hasta una medida limitada, el inverso no es cierto. Los grupos incapaces de producir vástagos fértiles con cada uno se les considera universalmente especies distintas, y no meramente “razas” diferentes de la misma especie.[cita requerida]
Aunque este intento de precisión conceptual se extendió con muchos biólogos, especialmente los zoólogos, los científicos evolucionistas lo han criticado en un número de frentes.[cita requerida]
El rechazo de raza y el auge de población y cline
A principio del siglo XX, los antropólogos cuestionaron, para luego abandonar, la afirmación de que razas biológicamente distintas son isomórficas respecto a diferentes grupos lingüísticos, culturales y sociales. Entonces el desarrollo de la genética de poblaciones llevó a algunos estudiosos de la antropología y la biología, simpatizantes con el evolucionismo, a cuestionarse sobre la validez de raza como concepto científico que describiera un fenómeno real y objetivo. Quienes vinieron a rechazar la validez del concepto raza, lo hicieron por cuatro razones: por evidencia empírica, por definición, por disponibilidad de conceptos alternativos, y por cuestiones éticas (Lieberman & Byrne, 1993).
Los primeros en cuestionar el concepto de raza en el terreno empírico fueron los antropólogos Franz Boas —que demostró la plasticidad fenotípica debida a factores mediambientales (Boas, 1912)— y Ashley Montagu (1941, 1942), que se apoyaba en datos producidos por la genética. Los zoólogos Edward O. Wilson y W. Brown también pusieron en tela de juicio el concepto desde la perspectiva de la clasificación científica general, y posteriormente rechazaron la afirmación que “raza” era equivalente a “subespecie” (Wilson and Brown 1953). En Race and History (Unesco, 1952), Claude Lévi-Strauss impuso las propuestas provenientes del relativismo cultural, con la famosa metáfora de la culturas como trenes cruzándose unos con otros en diferentes direcciones, cada uno viendo a los otros inmóviles mientras iban avanzando.
Una de las innovaciones cruciales para la nueva concepción de las variaciones genotípicas y fenotípicas fue la observación del antropólogo C. Loring Brace. Sostenía que dichas variaciones, en tanto que están afectadas por la selección natural, la migración, o la deriva genética, se encuentran distribuidas en gradaciones geográficas llamadas “clines” (Brace 1964). Este punto llamó la atención en un problema común en las descripciones de las razas basadas en los fenotipos —consideraciones sobre la textura del pelo y el color de piel—: ignoran una gran cantidad de otras similitudes y diferencias (por ejemplo, el tipo de sangre) que no se correlacionan altamente con los demarcadores de la raza. Así, la conclusión del antropólogo Frank Livingstone que, desde que los clines cruzan fronteras raciales, “no hay razas, solo clines” (Livingstone, 1962: 279). En 1964, los biólogos Paul Ehrlich y Holm apuntaron a casos donde dos o más clines están distribuidos discordantemente —por ejemplo, la melanina se distribuye siguiendo un patrón decreciente desde el norte del ecuador al sur; las frecuencias para el haplotipo para la hemoglobina beta-S, por otro lado, irradian de puntos geográficos específicos en África (Ehrlich & Holm, 1964). Como los antropólogos Leonard Lieberman y Fatimah Linda Jackson observan, “Los patrones discordantes de heterogeneidad falsifican cualquier descripción de una población como si esta fuera genotípicamente o incluso fenotípicamente homogénea” (Lieverman & Jackson, 1995).
Finalmente, el genetista Richard Lewontin, al observar que el 85 por ciento de las variaciones humanas ocurren dentro de la población, y no entre las poblaciones, argumentó que ni “raza” ni “subespecie” eran formas apropiadas o útiles de describir las poblaciones (Lewontin, 1973). Esta visión es descrita por sus detractores como la falacia de Lewontin. Algunos investigadores dicen que la variación entre grupos raciales (medida por la estructura estadística de población de Sewall Wright (FST) cuenta tan poco como el 5 % de la variación genética humana². Sin embargo, debido a las limitaciones técnicas de FST, muchos genetistas hoy día creen que los valores bajos FST no invalidan la posibilidad de que puedan existir diferentes razas humanas (Edwards, 2003). Por otra parte, el consenso científico actual dice que la raza es una construcción social no basada en nuestra realidad biológica. Neomarxistas como David Harvey (1982, 1984, 1992) añaden que se usa en cambio para debilitar las diferencias entre clases.
Estas cuestiones empíricas del concepto de raza forzaron a las ciencias evolucionistas a reconsiderar su definición de raza. William Boyd, antropólogo de mediados de siglo XX, definió raza como:
Una población que difiere significativamente de otras poblaciones respecto a la frecuencia de uno o más genes que posee. Es una cuestión arbitraria cuales, y cuantos, gene loci escogemos para considerar una “constelación” significativa.Boyd:1950
Lieberman y Jackson (1994) han apuntado que “la debilidad de este argumento es que si un gen puede distinguir razas entonces el número de razas es tan numeroso como el número de parejas humanas reproduciéndose”. Además, el antropólogo Stephen Molnar ha sugerido que la discordancia de los clines inevitablemente resulta en una multiplicación de razas que inutiliza la riqueza teórica del concepto (Molnar, 1992).
Junto con los problemas empíricos y conceptuales de la "raza" después de la Segunda Guerra Mundial, los científicos sociales evolucionistas fueron sumamente conscientes de la forma en que las creencias sobre las razas habían servido para “legitimar” la discriminación, el apartheid, la esclavitud, y el genocidio. Esta cuestión se volvió central en los años 1960, época del Movimiento por los Derechos Civiles en Estados Unidos y la aparición del anticolonialismo.
De cara a esta problemática, algunos científicos evolucionistas simplemente abandonaron el concepto de raza en favor de población. Lo que distingue población de las agrupaciones previas de humanos por raza es que la primera se refiere a una población que se reproduce —punto esencial para cálculos genéticos— y no a un taxón biológico. Otros científicos evolucionistas han abandonado el concepto de raza en favor de cline —concebido la frecuencia de un rasgo cambia a lo largo de un gradiente geográfico. Los conceptos de población y cline, sin embargo, no son mutuamente excluyentes y suelen ser empleados en el lenguaje científico de corte evolucionista.
Como corolario del rechazo de raza en el campo de las ciencias biológicas, varios científicos sociales han reemplazado el término por etnia, para referirse a los grupos propiamente identificados basados en las creencias compartidas como la religión, la nacionalidad, el territorio y otras categorías no necesariamente biológicas como la lengua y las costumbres. Además, entendieron que estas creencias y comportamientos compartidos no son constituyentes de una raza propiamente dicha sino que son construcciones sociales que no tienen bases objetivas en el reino natural o supernatural (Gordon, 1964).
Estudios recientes sobre la población brasileña indican que no hay relación entre genes y razas.
Resumen de las definiciones del término
| Concepto | Referencia | Definición |
|---|---|---|
| Esencialista | Earnest Albert Hooton (1926) | “Una gran división de la humanidad, caracterizada como grupo por compartir una cierta combinación de rasgos, que derivan de su ascendencia común, y constituyen un vago origen físico, normalmente más o menos ocultado por variaciones individuales, y adviertes mejor en una imagen compuesta.” |
| Taxonómico | Ernst Mayr (1969) | “Un conjunto de poblaciones fenotípicamente similares de una especie, que habitan una subdivisión geográfica del rango de una especie, y que difieren taxonómicamente de otras poblaciones de especies.” |
| Población | Theodosius Dobzhansky (1970) | “Las razas son poblaciones mendelianas genéticamente distintas. No son ni individuos ni genotipos particulares, consisten en individuos que difieren genéticamente entre ellos.” |
| Linaje | Alan Templeton (1998) | “Una subespecie (raza) es un linaje evolutivo distinto en una especie. Esta definición requiere que la subespecie sea genéticamente diferenciada debido a obstáculos en el intercambio genético que han persistido durante largos periodos de tiempo; esto es, las subespecies deben tener una continuidad histórica además de una diferenciación genética actual.” |
Variación física y color de piel
La distribución de muchos rasgos físicos se parece a la distribución de la variación genética dentro de poblaciones humanas entre estas (American Association of Physical Anthropologists 1996; Keita and Kittles 1997). Por ejemplo, ∼90 % de la variación en la forma de las cabezas humanas ocurre dentro de cada grupo humano, y ∼10 % separa los grupos, con una mayor variabilidad de las forma de la cabeza entre los individuos con antepasados africanos recientes (Relethford 2002).
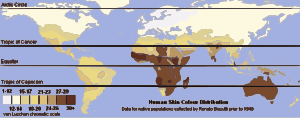
Una excepción prominente a la distribución común de las características físicas dentro de y entre grupos es el color de piel. Aproximadamente un 10 % de la variancia en el color de piel ocurre dentro de los grupos, y ~90 % ocurre entre grupos (Relethford 2002). Esta distribución del color de piel y su patrón geográfico —con gente con antepasados que vivieron predominantemente cerca del ecuador teniendo una piel más oscura que esos con antepasados que vivieron predominantemente en latitudes superiores— indican que este atributo ha estado bajo una fuerte presión selectiva. La piel más oscura parece estar fuertemente seleccionada para las regiones ecuatoriales para prevenir las quemaduras de sol, el cáncer de piel, la fotólisis de folate, y el daño a las glándulas sudoríparas (Sturm et al. 2001; Rees 2003). Una hipótesis destacada para la selección de un color de piel más claro en latitudes más altas es que permite al cuerpo formar mayores cantidades de vitamina D ya que en esas latitudes los rayos ultravioleta del sol son menos intensos , que ayuda a prevenir el raquitismo (Jablonski 2004). Las evidencias para esto incluyen el descubrimiento que una porción substancial de las diferencias de color de piel entre los europeos y los africanos reside en un único gen, SLC24A5 el alelo 111 de la treonina que fue encontrado entre el 98,7 y el 100 % entre varias muestras de europeos, mientras que la forma alanina-111 se encontró entre el 93 y 100 % de las muestras de africanos, asiáticos del este y amerindios (Lamason et al. 2005). Sin embargo, la hipótesis de la vitamina D no está universalmente aceptada (Aoki 2002), y un color de piel más claro en las latitudes más altas puede corresponder simplemente a la ausencia de la selección para la piel oscura (Harding et al. 2000). La melanina que sirve de pigmento, está localizado en la epidermis de la piel, y está basada en la expresión génica hereditaria.
Debido a que el color de piel ha estado bajo una fuerte presión selectiva, similares colores de piel pueden resultar de una adaptación convergente en vez de una relación genética. Los africanos subsaharianos, poblaciones tribales del sur de India, y los aborígenes australianos tienen una pigmentación de piel similar, pero genéticamente no son más similares que otros grupos ampliamente separados. Además, en algunas partes del mundo donde gente de diferentes regiones se han mezclado extensivamente, la conexión entre color de piel y ascendencia ha sido substancialmente debilitada (Parra et al. 2004). En Brasil, por ejemplo, el color de piel no está estrechamente asociado con el porcentaje de antepasados africanos recientes que una persona tiene, por estimaciones de un análisis de variantes genéticas que difieren en frecuencia entre los grupos continentales (Parra et al. 2003).
Una especulación considerable ha rodeado el posible valor adaptivo de otros rasgos físicos característicos de grupos, como la constelación de rasgos faciales observados en muchos asiáticos orientales y nororientales (Guthrie 1996). No obstante, cualquier característica física dada generalmente se encuentra en múltiples grupos (Lahr 1996), y demostrar que las presiones selectivas medioambientales formaron rasgos físicos específicos será difícil, ya que estos rasgos pueden haber resultado de la selección sexual de individuos con ciertas apariencias o de la deriva genética (Roseman 2004).
Variación genética humana
Cladística
Un árbol filogenético se obtiene normalmente de secuencias de ADN o proteínas de las poblaciones. A menudo las secuencias de ADN mitocondrial o cromosoma Y son usadas para estudiar antiguas poblaciones humanas. Estas fuentes de ADN simple de locus no se recombinan y son casi siempre heredadas de un único padre, con la única excepción conocida en mtDNA (Schwartz & Vissing, 2002). Los individuos de varios grupos continentales tienden a ser más similares entre ellos que con gente de otros continentes. El árbol tiene su raíz en el antepasado común de los chimpancés y los humanos, que se cree que se originó en África. La distancia horizontal corresponde a dos cosas:
- Distancia genética. Dado debajo del diagrama, la diferencia genética entre los humanos y los chimpancés es aproximadamente de un 2 por ciento, o 20 veces más grande que la variación entre los humanos modernos.
- Lejanía temporal del más reciente antepasado común. Las estimaciones aproximadas dadas en el diagrama superior, en millones de años. El más reciente antepasado mitocondrial común de los humanos modernos vivió hace aproximadamente apenas 25 000 años, los últimos antepasados comunes de los humanos y los chimpancés entre cuatro y siete millones de años atrás.
Los chimpancés y los humanos pertenecen a diferente género, indicado en rojo. La formación de especies y subespecies también está indicado, y la formación de “razas” está indicada en el rectángulo verde de la derecha (note que solo se da una aproximación representativa de la filogenia humana). Note que las distancias verticales no son significativas en esta representación.
Distribución de la variación
Una minuciosa descripción de las diferencias en los patrones de la variación genética entre los humanos y otras especies espera estudios genéticos adicionales de las poblaciones humanas y las especies no humanas. Pero los datos acumulados hasta ahora sugieren que la variación humana muestra varias características distintivas. Primero, comparado con muchas otras especies mamíferas, los humanos son genéticamente menos diversos— un hallazgo contraintuitivo, dada nuestra amplia población y distribución mundial (Li & Sadler, 1991; Kaessmann et al., 2001). Por ejemplo, las subspecies de chimpancé que viven solo en África central y occidental tienen niveles más altos de diversidad que los humanos (Ebersberger et al., 2002; Yu et al., 2003; Fischer et al., 2004).
Dos humanos aleatorios se espera que difieran aproximadamente en 1 de 1000 nucleótidos, mientras que dos chimpancés aleatorios difieren a 1 en 500 pares de nucleótidos. Sin embargo, con un génoma de aproximadamente 3 billones de nucleótidos, de media dos humanos difieren aproximadamente 3 millones de nucleótidos. La mayoría de estos polimorfismos de nucleótido simple (SNPs) son neutrales, pero algunos son funcionales e influencian las diferencias fenotípicas entre humanos. Se estima que sobre unos 10 millones de SNPs existen en las poblaciones humanas, donde el más raro alelo SNP tiene una frecuencia de al menos un 1%.
La distribución de las variantes dentro de las poblaciones humanas y entre ellas también difiere de la de muchas otras especies. Los detalles de esta distribución son imposibles de describir sucintamente por la dificultad de definir una “población”, la naturaleza clinal de la variación, y la heterogeneidad a través del genoma (Long & Kittles, 2003). En general, sin embargo, entre un 5% y un 15% de variación genética ocurre entre grandes grupos que viven en continentes diferentes, con el resto de la mayoría de la variación ocurriendo entre estos grupos (Lewontin, 1972; Jorde et al., 2000a; Hinds et al., 2005). Esta distribución de variación genética difiere de los patrones vistos en muchas otras especies mamíferas, por lo que los datos existentes sugieren una mayor diferenciación entre los grupos (Templeton, 1998; Kittles & Weiss, 2003).
Nuestra historia como especie también ha dejado señales genéticas en poblaciones regionales. Por ejemplo, además de tener niveles mayores de diversidad genética, las poblaciones en África tienden a tener menores cantidades de desequilibrio de ligamiento que otras poblaciones fuera de África, en parte por la mayor cantidad de poblaciones humanas en África durante el curso de la historia humana y en parte porque el número de humanos modernos que dejaron África para colonizar el resto del mundo parece haber sido relativamente bajo (Gabriel et al., 2002). En cambio, las poblaciones que han experimentado reducciones o expansiones dramáticas de tamaño en el pasado y las poblaciones formadas por la mezcla de grupos antepasados separados previamente pueden tener inusualmente altos niveles de desequilibrio de enlace (Nordborg & Tavare, 2002).
En el campo de la genética de poblaciones, se cree que la distribución de polimorfismos neutrales entre los humanos contemporáneos refleja la historia demográfica humana. Se cree que los humanos pasaron a través de un cuello de botella poblacional antes de una rápida expansión coincidiendo con las migraciones de África llevando a una divergencia africano-eurasiática cerca de 100.000 años atrás (ca. 5000 generaciones), seguido de una divergencia europeo-asiática hace unos 40.000 años (ca. 2.000 generaciones).
La rápida expansión de una previamente pequeña población tiene dos importantes efectos en la distribución de la variación genética. Primero, el llamado efecto fundador ocurre cuando las poblaciones fundadoras traen solo un subconjunto de la variación genética de su población ancestral. Segundo, a medida que los fundadores se separan más geográficamente, la probabilidad que dos individuos de diferentes poblaciones fundadoras se aparearan se convierte en menor. El efecto de este emparejamiento selectivo es reducir el flujo genético entre grupos geográficos, y aumentar la distancia genética entre los grupos. La expansión de los humanos desde África afectó a la distribución de la variación genética de otras dos maneras. Primero, las poblaciones más pequeñas (fundadoras) experiencian mayor deriva genética por la fluctuaciones aumentadas en los polimorfismos neutrales. Segundo, los nuevos polimorfismos que aparecen en un grupo fueron menos probables de ser transmitidos a otros grupos ya que el flujo genético era restringido.
Muchos otros factores geográficos, climáticos, e históricos han contribuido a los patrones de la variación genética humana observada en el mundo hoy en día. Por ejemplo, procesos de población asociados con la colonización, periodos de aislamiento geográfico, endogamía reforzada socialmente, y la selección natural todos han afectado a las frecuencias de alelos en ciertas poblaciones (Jorde et al., 2000b; Bamshad & Wooding, 2003). En general, sin embargo, la recencia de nuestra ascendencia común y continuo flujo genético entre grupos humanos ha limitado la diferenciación genética en nuestra especie.
Subestructura en la población humana
Los nuevos datos sobre la variación genética humana ha reencendido el debate en torno a la raza. La mayoría de la controversia rodea la cuestión de cómo interpretar estos nuevos datos, y si las conclusiones basadas en los datos existentes son sensatos. Una gran mayoría de investigadores apoyan la opinión que los grupos continentales no constituyen subespecies diferentes. Sin embargo, otros investigadores siguen debatiendo si los linajes evolucionistas deben correctamente llamarse “razas”. Estas cuestiones son particularmente urgentes para la biomedicina, donde una raza propiamente descrita es a menudo usado como un indicador de ascendencia.
La evidencia biológica moderna del libro de texto sobre antropología Human Species (2003) contradice las teorías anteriores de que los grupos estaban más genéticamente relacionados con otros grupos. Los humanos están todos relacionados. La humanidad dividida en las ramas africanas y eurásiatica/oceánica. Las ramas eurásiaticas y oceánicas son producto de este origen común. La rama eurasiática se separó en los amerindios y la rama mayor asiática del este. La rama mayor asiática del este se dividió a su vez en rusos orientales y asiáticos del este. La rama oceánica se dividió en asiáticos de sureste e isleños del Pacífico. Según Human Species (2003), los asiáticos del este son generalmente más similares genéticamente a los asiáticos del sur que a los asiáticos del sudeste, porque el Extremo Oriente y el subcontinente indio son miembros de la rama eurasiática mientras que los asiáticos del sudeste (incluyendo a los chinos del sur) son miembros de la rama oceánica. De modo más interesante, los asiáticos tienen clusters genéticos muy locales dentro de estas regiones, significando que diferentes grupos étnicos asiáticos no se han entrecruzado históricamente los unos con los otros. Ejemplos de clusters genéticos localizados incluyen Japón, Corea, Mongolia y China que forman clusters genéticos separados unos de otros.
Aunque las diferencias genéticas entre grupos humanos son relativamente pequeñas, estas diferencias en ciertos genes (como duffy, ABCC11, SLC24A5, llamados marcadores informativos de la ascendencia) pueden utilizarse, no obstante, para situar fiablemente muchos individuos dentro de un grupo o raza amplia basada geográficamente. Por ejemplo, análisis con ordenador de miles de muestras de locis polimórficos abundantes en las poblaciones globalmente distribuidas han revelado la existencia de agrupaciones genéticas que aproximadamente se asocian con grupos que históricamente han ocupado amplias regiones continentales y subcontinentales (Rosenberg et al., 2002; Bamshad et al., 2003).
Algunos comentaristas han expuesto que estos patrones de variación proporcionan una justificación biológica para el uso de las categorías raciales tradicionales. Argumentan que las agrupaciones continentales corresponden aproximadamente con la división de los seres humanos en africanos subsaharianos; europeos, asiáticos occidentales, y africanos del norte; asiáticos orientales; polinesios y otros habitantes de Oceanía; y amerindios (Risch et al., 2002). Otros observadores discrepan, diciendo que los mismos datos recortan las nociones tradicionales de grupos raciales (King & Motulsky, 2002; Calafell, 2003; Tishkoff & Kidd, 2004). Apuntan, por ejemplo, que las grandes poblaciones consideradas razas o subgrupos dentro de las razas no necesariamente forman sus propios clusters. Así, muestras tomadas de India y Pakistán los afilian con europeos o asiáticos orientales en vez de separarlos en clusters distintos.
Además, porque la variación genética humana es clinal, muchos individuos se afilian con dos o más grupos continentales. Así, la “ascendencia biogeográfica” basada genéticamente asignada a cualquier persona dada generalmente será ampliamente distribuida y estará acompañada de incertidumbres considerables (Pfaff et al., 2004).
En muchas partes del mundo, los grupos se han mezclado de tal manera que muchos individuos tienen antepasados relativamente recientes de regiones extensamente separadas. Aunque los análisis genéticos de grandes números de loci pueden producir estimaciones del porcentaje de los antepasdos de una persona viniendo de varias poblaciones continentales (Shriver et al., 2003; Bamshad et al., 2004), estas estimaciones pueden suponer una falsa distinción de las poblaciones parentales, ya que los grupos humanos han intercambiado apareamientos desde escalas locales a continentales durante toda la historia (Cavalli-Sforza et al., 1994; Hoerder, 2002). Incluso con grandes números de marcas, la información para estimar las proporciones de la mezcla de los individuos o grupos es limitada, y las estimaciones típicamente tendrán amplios CIs (Pfaff et al., 2004).
Para los linajes genéticos paternos y maternos del mundo, véase: [6] y [7]
Orígenes y evolución de los humanos modernos
Cualquier modelo biológico de raza debe servir para explicar el desarrollo de las diferencias raciales durante la evolución humana. No obstante, la mayor parte del siglo XX los antropólogos contaron con un registro fósil incompleto para reconstruir el proceso que ha llevado a la aparición del Homo sapiens. Sus modelos rara vez proporcionaron una base firme para sostener sus inferencias sobre el origen de las razas. En contraste, las investigaciones modernas en biología molecular han proporcionado a los científicos evolucionistas con un tipo de datos completamente nuevo, que se añaden para mejorar el conocimiento de nuestro pasado.
A través de la historia, ha habido un debate considerable entre los antropólogos sobre los orígenes del Homo sapiens. Un millón de años atrás el Homo erectus migró de África a Europa y Asia. El debate gira en torno a la posibilidad de que H. erectus haya evolucionado a H. sapiens más o menos simultáneamente en África, Europa y Asia; o si el H. sapiens evolucionó solo en África y finalmente sustituyó al Homo erectus, al Homo neanderthal y al Homínido de Denisova en Europa y Asia. Cada modelo sugiere diferentes escenarios posibles para la evolución de las distintas razas. Sin embargo la evidencia genética actual apoya y refuerza la hipótesis de que el H. sapiens surgió en África y de ahí conquistó el mundo; esta es actualmente la opinión mayoritaria.
Respecto a la genética de los diferentes fenotipos, las últimas investigaciones indican que al salir el H. sapiens de África, existió posteriormente una hibridación con otras especies “razas” homínidas más antiguas, tales como el Homo neanderthalensis (de un 1 % a un 4 % de genes neandertales por persona, y de un 20 % en su sumatoria del porcentaje total de genes dentro del genoma de la población actual, principalmente en Europa); e igualmente hibridación con el Homínido de Denisova (la población local que vive actualmente en Papúa Nueva Guinea, en el Sudeste Asiático, le debe al menos el 3 % de su genoma por persona a los Homínidos de Denisova). Esta hibridación habría traído como consecuencia que en la “heterosis” humana producida naturalmente, para el caso del genoma neandertal heredado, produjo cambios en el fenotipo; como ejemplo fenotipos de la piel, mediante la inclusión de genes implicados en las adaptaciones ambientales a climas de Europa y Asia. Así esta hibridación con diferentes especies (“razas”) humanas antiguas, habría igualmente tenido un papel importante en los fenotipos asociados a algunas “razas” en el ser humano actual
Interpretaciones sociales
Los historiadores, antropólogos y científicos sociales a menudo describen las razas humanas como una construcción social, prefiriendo en vez del término población, al cual se le puede dar una clara definición operacional. Incluso esos que rechazan el concepto formal de raza, sin embargo, siguen usando la palabra raza en el habla diaria. Esto puede ser una cuestión de semántica, o un efecto de un significado cultural de raza subyacente en sociedades racistas. A pesar del nombre, un concepto de trabajo de agrupación de subespecies puede ser útil, porque en la ausencia de tests genéticos baratos y extendidos, varias mutaciones genéticas relacionadas con la raza (fibrosis quística, intolerancia a la lactosa, enfermedad de Tay-Sachs y anemia de células falciformes) son difíciles de tratar sin recurrir a una categoría entre “individuo” y “especie”.
En el habla diaria, raza a menudo describe poblaciones mejor definidas como etnias, con frecuencia conduciendo a discrepancias entre visiones científicas sobre la raza y el uso popular del término. Por ejemplo en muchas partes de los Estados Unidos, categorías como hispano o latino se ven como que constituyen una raza, aunque otros ven hispano como una agrupación lingüística y cultural que viene de una variedad de procedencias. En Europa, una distinción, sugerir que europeos del Sur no son europeos o blancos, parecerá extraño como mínimo o posiblemente incluso insultante. En los Estados Unidos, en lo que es referido como la one-drop rule (Regla de una gota), el término negro subsume gente con un amplio rango de ascendencias bajo una etiqueta, aunque muchos de los que son llamados negros pueden ser descritos más precisamente como blancos a través de un simple método antropológico o taxonómico. En gran parte de Europa grupos como los gitanos y los indios son definidos comúnmente como racialmente distintos de los europeos “blancos”, aunque estos grupos pueden ser considerados “caucásicos” por los antiguos métodos antropológicos físicos que emplean medidas finitas de nariz y la estructura del cráneo como la forma estándar de clasificación racial.
Algunos argumentan que es preferible cuando se consideran las relaciones biológicas el pensar en términos de poblaciones, y cuando se consideran las relaciones culturales pensar en términos de origen étnico, en vez de raza. En lugar de clasificar la gente en un “grupo”, digamos “caucásicos” o europeos tienes británicos, franceses, alemanes, nórdicos, eslavos del oeste y celtas más que tener un término implicando un (posible) grupo de ascendencia en el Cáucaso que es definitivamente demasiado distante para cualquier consideración real, y además alcanza a grupos incluyendo eslavos del este, gitanos, así como georgianos, y otros que difieren notablemente, ambos en cultura, y en una medida digna de mención en la apariencia física, de los mencionados grupos étnicos. Puede haber tanta diferencia entre dos etnias agrupadas en una única “raza” como puede haber entre etnias agrupadas (a menudo arbitrariamente) en otra “raza”.
Estos desarrollos tuvieron importantes consecuencias. Por ejemplo, algunos científicos desarrollaron la noción de “población” para tomarle el lugar a raza. Esta substitución no es simplemente una cuestión de intercambiar una palabra por otra. Las poblaciones son, en un sentido, simplemente clusters estadísticos que surgen de la elección de variables de interés; no hay un conjunto de variables preferidas. La visión “poblacionista” no niega que hay diferencias físicas entre gentes; simplemente reclama que las concepciones históricas de “raza” no son particularmente útiles en la importancia de estas diferencias científicamente.
Desde los años 1960, algunos antropólogos y profesores de antropología han reconcebido “raza” como una categoría cultural o construcción social, en otras palabras, como una manera particular que alguna gente tiene de hablar de ellos y los otros. Por eso no puede ser un concepto analítico útil; mejor dicho, el uso del término “raza” por sí solo debe ser analizado. Además, ellos exponen que la biología no explicará por qué o cómo la gente usa la idea de raza: la historia y las relaciones sociales lo harán.
Diversidad étnica y racial en el Nuevo Mundo postcolombino
Incluso mientras la idea de “raza” se estaba convirtiendo en un principio organizador poderoso en muchas sociedades, los defectos del concepto eran aparentes. En el Mundo Antiguo, la transición gradual en apariencias de un grupo a grupos adyacentes enfatizó que “una variedad de la humanidad pasa tan sensiblemente a otra, que no puedes marcar los límites entre ellas”, como Blumenbach observó en su escritos sobre la variación humana (Marks 1995, p. 54). En partes de las Américas, la situación era de alguna manera diferente. Los inmigrantes al Nuevo Mundo vinieron en cantidad de regiones ampliamente separadas del Viejo Mundo—Europa occidental y norte, África occidental, y, posteriormente, Ásia oriental y Europa del sur. En las Américas, las poblaciones inmigrantes comenzaron a mezclarse entre ellas y con los habitantes indígenas del continente. En los Estados Unidos, por ejemplo, la mayoría de gente que se identifica como afrodescendiente tiene algunos antepasados europeos—en un análisis de las marcas genéticas que tienen diferentes frecuencias entre los continentes, la ascendencia europea osciló ente un estimado 7 % para una muestra de jamaicanos a un ∼23 % para una muestra de afroamericanos de Nueva Orleans (Parra et al. 1998). Similarmente, mucha gente que se identifica como euroamericano tiene algunos antepasados africanos o americanos nativos, bien a través de matrimonios abiertamente interraciales o a través de la inclusión gradual de gente con ascendencia mezclada en la mayoría de la población. En una encuesta de unos estudiantes de college que se identificaron como blancos en una universidad del noroeste de Estados Unidos, ∼30 % fueron estimados de tener <90 % de ascendencia europea (Shriver et al. 2003).
En los Estados Unidos, se desarrollaron durante un tiempo unas convenciones sociales y legales que forzaron a los individuos de ascendencia mixta en categorías raciales simplificadas (Gossett 1997). Un ejemplo es la one-drop rule implementada en algunas leyes de estados que trataron a cualquiera con un único antepasado conocido afrodescendiente como negro (Davis 2001). Los censos de cada década realizados desde 1790 en los Estados Unidos también crearon un incentivo para establecer categorías raciales y meter gente en esas categorías (Nobles 2000). En otros países en las Américas donde la mezcla entre grupos era más extensiva, las categorías sociales han tendido a ser más numerosas y fluidas, con gente moviéndose de una categoría a otra sobre la base de la combinación del estado socioeconómico, la clase social, la ascendencia, y la apariencia (Mörner 1967).
Los esfuerzos hechos para ordenar la creciente población mestiza de los Estados Unidos en categorías discretas generó muchas dificultades (Spickard 1992). Por los estándares usados en los censos pasados, muchos millones de niños nacidos en los Estados Unidos han pertenecido a diferentes razas de las que tienen uno de sus padres biológicos. Los esfuerzos de seguirle la pista al mestizaje entre grupos llevó a la proliferación de categorías (como “mulato” y octoroon) y blood quantum distinciones que se convirtieron crecientemente desatadas de la ascendencia. La identidad racial de una persona puede cambiar durante el tiempo, y la raza autoatribuida puede diferir de la raza asignada (Kressin et al. 2003). Hasta el censo del 2000, los latinos debían identificarse como una raza separada a pesar de la larga historia de mestizaje en Latinoamérica; en parte como resultado de la confusión generada por la distinción, el 42 % de los latinos demandados en el censo del 2000 ignoraron las categorías especificadas y marcaron “alguna otra raza” (Mays et al. 2003).
La raza en los Estados Unidos
En los Estados Unidos desde su historia reciente, los americanos nativos, afroamericanos y euroamericanos fueron clasificados como pertenecientes a diferentes razas. Durante cerca de tres siglos, los criterios para ser miembro de estos grupos eran similares, comprendiendo la apariencia de la persona, su fracción de ascendencia no blanca conocida, y su círculo social.2 Pero los criterios para ser miembro de estas razas divergieron a finales del siglo XIX. Durante la Reconstrucción, números crecientes de americanos comenzaron a considerar cualquiera con “una gota” de “sangre” negra ser negro.3 A principios del siglo XX, esta noción de negreza invisible fue hecha estatutoria en muchos estados y ampliamente adoptada a escala nacional.4 En cambio, los amerindios continúan siendo definidos por un cierto porcentaje de “sangre india” (llamada blood quantum) debido en gran parte a éticas de la esclavitud americanas. Finalmente, durante el siglo pasado, para ser blanco uno debía tener ascendencia blanca “pura”.(Los americanos con aspecto completamente europeo de ascendencia hispana o árabe son excepciones en ser vistos como blancos por la mayoría de los americanos a pesar de conocidos rastros de ascendencia africana.)
La diferencia entre como las identidades americano nativo y negro están definidas hoy en día (blood quantum versus one-drop) ha pedido una explicación. Según antropólogos como Gerald Sider, el objetivo de estas designaciones raciales era concentrar el poder, la riqueza, los privilegios y las tierras en las manos de los blancos en una sociedad de hegemonía blanca y privilegios para los blancos (Sider 1996; véase también Fields 1990). Las diferencias tienen poco que ver con la biología y bastante más que ver con la historia del racismo y formas específicas de supremacía blanca (el orden del día social, geopolítico y económico de los blancos dominantes vis-à-vis subordinan los negros y los americanos nativos) especialmente los diferentes roles que los negros y los amerindios ocuparon en el siglo XIX dominado por los blancos en América. La teoría sugiere que la definición de blood quantum de la identidad de los americanos nativos permitió a los blancos comprar tierras amerindias, mientras que la one-drop rule de la identidad negra permitió a los blancos preservar su fuerza de trabajo en la agricultura. El contraste presumiblemente surgió porque a medida que la gente era transportada de lejos de su tierra y sus ataduras parentescas a otros continentes, el trabajo de los negros era relativamente fácil de controlar, de esta manera reduciendo los negros a valiosas mercancías como trabajadores en la agricultura. En cambio, el trabajo de los amerindios era más difícil de controlar; además, los amerindios ocuparon grandes territorios que se convirtieron en valiosos como tierras para la agricultura, especialmente con la invención de nuevas tecnologías como el ferrocarril; así, la definición blood quantum elevó la adquisición de tierras amerindias por los blancos en una doctrina del Destino Manifiesto que los expuso a la marginación y múltiples episodios de campañas de exterminación localizadas.
La política económica de la raza tuvo diferentes consecuencias para los descendientes de los americanos aborígenes y los esclavos africanos. El siglo XIX la regla de blood quantum significó que era relativamente más fácil para una persona de ascendencia mestiza euroamerindia ser aceptada como blanca. Los vástagos de solo unas pocas generaciones de endogamia entre amerindios y blancos probablemente no habrían sido considerados amerindio —al menos no en el sentido legal. Los amerindios podían tener un tratado de derechos sobre la tierra pero porque un individuo con un bisabuelo amerindio no seguía siendo clasificado como amerindio, perdieron cualquier reclamación legal de las tierras amerindias. Según la teoría, esto permitió a los blancos adquirir tierras amerindias. La ironía es que los mismos individuos que se les pudo denegar el prestigio legal porque eran “demasiado blancos” para pedir derechos de propiedad, podíam seguir siendo suficientemente amerindios para considarse como “razas”, estigmatizado por su ascendencia americana nativa.
La one-drop rule del siglo XX, por otro lado, hizo relativamente difícil para cualquiera de ascendencia negra conocida el ser aceptado como blanco. El hijo de un afrodescendiente cosechador y una persona blanca era considerado negro. Y, significante en términos de la economía de las cosechas, esa persona probablemente también sería un cosechador, de manera que se añadía a la fuerza de trabajo del empleador.
En resumen, esta teoría sugiere que en la economía del siglo XX que se benefició de los cosechadores, era útil tener el mayor número de negros posible. Al contrario, en el siglo XIX la nación se inclinó a una expansión hacia el oeste, era aventajoso disminuir los números de estos que podían reclamar el título de las tierras amerindias por simplemente definirlas fuera de la existencia.
Debe mencionarse, sin embargo, que aunque algunos eruditos del periodo de Jim Crow están de acuerdo que la noción de negreza invisible del siglo XX cambió la línea de color en la dirección de la palidez, de ese modo hinchando la fuerza de trabajo en respuesta a la gran migración hacia el norte de los negros del sur, otros (Joel Williamson, C. Vann Woodward, George M. Fredrickson, Stetson Kennedy) ven la one-drop rule como una simple consecuencia de la necesidad de definir blanqueza como ser puro, por tanto justificando la opresión de los blancos sobre los negros. En cualquier evento, durante los siglos que los blancos ejercieron el poder sobre los negros y los amerindios y creían ampliamente en su superioridad inherente sobre la gente de color, no es una coincidencia que el grupo racial más duro donde probar la pertenecía eran los blancos.
El término “hispano” surgió en el siglo XX con el crecimiento de la migración de trabajadores de países de habla hispana hacia los Estados Unidos; por tanto incluyen gente que ha sido considerada racialmente distinta (negros, blancos, amerindios) en sus países de nacimiento. Hoy en día, la palabra “latino” se usa a menudo como un sinónimo para “hispano” (la identificación de los países de lengua hispana en las Américas como “Latinoamérica” fue primero fomentado por los partidarios de Maximiliano como emperador de México en 1864. Maximiliano fue instaurado por el emperador francés Napoleón III como una manera de extender la influencia francesa en las Américas; ya que el francés y el español derivan ambos del latín, los franceses identificaron a los hablantes en español como “latinos” para enfatizar un parentesco ficticio con los franceses, y con el deseo —no cumplido— de legitimar a Maximiliano. En contraste con “latino”, “anglo” se usa hoy en día de una manera similar para referirse a los descendientes de los colonos británicos, y los valores y prácticas derivados de la cultura británica.
La raza en Brasil
Comparado con el siglo XIX de Estados Unidos, en el siglo XX Brasil estaba caracterizado por una relativa ausencia de grupos raciales firmemente definidos. Este patrón refleja una historia diferente y unas diferentes relaciones sociales. Básicamente, la raza en Brasil fue “biologizada”, pero de una manera que reconocía la diferencia entre la ascendencia (que determina el genotipo) y las diferencias fenotípicas. Allí, la identidad racial no fue gobernada por una regla rígida de descendencia. Un hijo de brasileños no era nunca automáticamente identificado con el tipo de raza de uno o ambos padres, como tampoco había solo dos categorías de las que escoger. Más de una docena de categorías raciales serían reconocidas en conformidad con las combinaciones del color y textura del pelo, el color del ojo, y el color de la piel. Estos tipos se clasifican en cualquier otra como los colores del espectro, y ninguna categoría se encuentra significativamente aislada del resto. Esto es, la raza referida a la apariencia y no a la herencia.
A través de este sistema de identificación racial, padres e hijos, e incluso hermanos y hermanas estuvieron frecuentemente aceptadas como representantes de tipos raciales opuestos. En un pueblo pescador en el estado de Bahía, un investigador mostró a 100 personas imágenes de tres hermanas y se les preguntó identificar las razas de cada una. En solo seis respuestas las hermanas fueron identificadas con el mismo término racial. Catorce respuestas usaron un término diferente para cada hermana. En otro experimento nueve retratos fueron mostrados a cien personas. Cuarenta tipos raciales diferentes fueron obtenidos. Se encontró, además, que dado un brasileño, éste puede llamarse por hasta trece términos diferentes por otros miembros de la comunidad. Estos términos están dispersados a través prácticamente del espectro entero de los tipos raciales teóricos. Una consecuencia adicional de la ausencia de una regla de descendencia fue que los brasileños aparentemente no solo no estaban de acuerdo con la identidad racial de individuos específicos, sino que también parecieron estar en desacuerdo sobre el significado abstracto de los términos raciales definidos por palabras y frases. Por ejemplo, el 40 % de una muestra lo clasificó como moreno claro (persona “clara” de ascendencia primariamente europea con pelo oscuro) como un tipo más claro que mulato claro (persona “clara” de ascendencia mezclada europea y africana), mientras el 60 % invirtió este orden. Una nota adicional de confusión es que una persona puede emplear diferentes términos raciales para describir la misma persona durante un corto lapso de tiempo. La elección de cuál descripción racial usar puede variar según las relaciones personales y el carácter de los individuos involucrados. El censo brasileño lista la raza de uno según la preferencia de la persona entrevistada. Como consecuencia, cientos de razas aparecieron en los resultados del censo, oscilando entre azul (que es más negro que el negro típico) a verde (que es más blanco que el blanco típico).
Así, aunque la identificación de una persona por la raza es mucho más fluida y flexible en Brasil que en los Estados Unidos, sigue habiendo estereotipos raciales y prejuicios. Los rasgos africanos son considerados menos deseables; los negros han sido considerados socialmente inferiores, y los blancos superiores. Estos valores de supremacía blanca parecen ser un legado obvio de la colonización europea y el sistema de plantación basado en esclavos. La complejidad de las clasificaciones raciales en Brasil es reflejo en gran medida del mestizaje en la sociedad brasileña, que permanece altamente, pero no estrictamente, estratificada junto con las línea de color. De ahora en adelante, el mito brasileño de un país perfectamente “postracista”, compuesto de la “raza cósmica” celebrada en 1925 por José Vasconcelos, debe ser tomado con cautela, como el sociólogo Gilberto Freyre demostró en 1933 en Casa Grande e Senzala.
Usos prácticos de la “raza”
La raza en la política y la ética
Durante la Ilustración, las clasificaciones raciales se usaron para justificar la esclavitud de esos estimados de ser “inferiores”, las razas no blancas, y así teóricamente mejor encajados para toda la vida de trabajo duro bajo la supervisión blanca. Estas clasificaciones hicieron que la distancia entre razas pareciera casi tan extensa como entre especies, facilitando preguntas inquietantes sobre lo apropiado de este tratamiento de los humanos. La práctica era durante aquel tiempo generalmente aceptada por las comunidad científica.
Essai sur l'inégalité des races humaines (1853-1855) de Arthur de Gobineau fue uno de los hitos en el nuevo discurso racista, junto con la “antroposociología” de Vacher de Lapouge y Johann Gottfried von Herder, que aplicó la raza a la teoría nacionalista para desarrollar un nacionalismo étnico militante. Ellos propusieron la existencia histórica de razas nacionales como los alemanes y los franceses, bifurcandóse de razas fundamentales supuestas de haber existido durante milenios, como la raza aria, y creyeron que las fronteras políticas debían reflejar estas supuestas razas.
Posteriormente, uno de los dichos favoritos de Hitler fue, “La política es biología aplicada”. Las ideas de Hitler de pureza racial llevaron a atrocidades tan solo superadas por el genocidio hacia el pueblo ucraniano durante la dictadura de José Stalin, en el cual más de 7 millones de ucranianos (niños, mujeres y ancianos en su mayoría fueron) murieron de hambre, o el genocidio Chino hacia el Tíbet en donde desde 1950 entre 1 y 2 millones de tibetanos han sido asesinados: torturados, violados, mutilados, apaleados hasta morir, envenenados, etc.. Desde entonces, la limpieza étnica ha ocurrido en los Balcanes y Ruanda. En un sentido, la limpieza étnica es otro nombre para la guerra tribal y el homicidio de masas que ha afligido a la sociedad humana durante años, pero estos crímenes parecen ganar intensidad cuando creen haber sido sancionados científicamente.
La desigualdad racial ha sido una preocupación de los políticos y legisladores de los Estados Unidos desde la fundación del país. En el siglo XIX la mayoría de los americanos blancos (incluyendo los abolicionistas) explicaron la desigualdad racial como una consecuencia inevitable de las diferencias biológicas. Desde mediados del siglo XX, los líderes políticos y cívicos así como los científicos han debatido en que medida la desigualdad racial es cultural en origen. Algunos sostienen que estas desigualdades actuales entre los negros y los blancos son ante todo culturales e históricas, el resultado del racismo anterior, la esclavitud y la segregación, y pueden ser reparadas a través de programas como la acción afirmativa y el Head Start. Otros trabajan para reducir los pagos de impuestos de programas de recuperación para las minorías. Han basado su apoyo en datos de tests de aptitud, que según ellos, muestran que las diferencias raciales en las capacidades son de origen biológico y no pueden ser nivelados incluso por intensivos esfuerzos pedagógicos. En políticas electorales, muchas más minorías étnicas han ganado importantes cargos en las naciones occidentales que en tiempos anteriores, aunque los cargos más altos tienden a permanecer en las manos de los blancos.
En su famosa Carta desde la cárcel de Birmingham, el Rev. Dr. Martin Luther King observó:
- La historia es el relato largo y trágico del hecho de que rara vez los grupos privilegiados renuncian a sus privilegios voluntariamente. Los individuos pueden ver la luz moral y voluntariamente dejar su postura injusta; pero como Reinhold Niebuhr nos recordó, los grupos son más inmorales que los individuos.
La esperanza del Dr. King, expresada en su discurso Yo tengo un sueño, fue que la lucha por los derechos humanos un día producirá una sociedad donde la gente no será “juzgada por el color de su piel, sino por el contentido de su carácter”.
Por la identificación del concepto de raza con la opresión política, muchos científicos naturales y sociales hoy en día son cautelosos al usar raza para describir la variación humana. Algunos, sin embargo, sostienen que la raza es no obstante de continua utilidad y validez en la investigación científica. La ciencia y la política frecuentemente toman lados opuestos en debates relacionados con la inteligencia humana y la biomedicina.
La raza y la inteligencia
Los investigadores han informado de diferencias significativas en la puntuación media de Cociente intelectual en los test de varios grupos étnicos. La interpretación y causas de estas diferencias han generado varias hipótesis, algunas de ellas son altamente controvertidas. Algunos investigadores, como Arthur Jensen, Richard Herrnstein, y Richard Lynn han argumentado que estas diferencias son como mínimo parcialmente genéticas sin embargo este punto de vista esta bastante desacreditado y ha sido ampliamente rechazado por medios científicos y no científicos y se puede decir que es solo esposado por una minoría extrema. Otros, como Stephen Jay Gould y Richard Lewontin, consideran que las categorías como “raza” e “inteligencia” son construcciones culturales que vuelven este tipo de investigaciones científicamente imperfectas. Algunos otros rehúyen una posición definida, por ejemplo Thomas Sowell, evitan la cuestión de los orígenes de la categorización y buscan explicar la distancia en la puntuación de los tests en términos de diferencias sociales que afectan en cuánto las capacidades innatas que cualquier persona individual puede conseguir. Estudios recientes indican una débil relación entre la inteligencia y los genes; lo cual descarta la idea de razas o fenotipos con una inteligencia determinada. Otros estudios indican que no hay una conexión estrecha entre personalidad, carácter y genética, sugiriendo una relación débil entre la psicología y los genes. Finalmente, la plasticidad del cerebro humano permite modificaciones voluntarias del cociente intelectual mediante diversos métodos como la meditación.
La raza en la biomedicina
Hay un debate activo entre los investigadores biomédicos sobre el significado y la importancia de la raza en su investigación. El ímpetu principal para considerar la raza en la investigación biomédica es la posibilidad de mejorar la prevención y el tratamiento de enfermedades predeciendo los factores difíciles de establecer sobre la base de las características más fácilmente establecidas. Los ejemplos más conocidos de desórdenes determinados genéticamente que varían en incidencia entre grupos étnicos serían la anemia de células falciformes y la talasemia entre los negros y las poblaciones mediterráneas y la enfermedad de Tay-Sachs entre gente de ascendencia Ashkenazi judía. Algunos temen que el uso de etiquetas raciales en la investigación biomédica corra el riego de exacerbar las disparidades de salud sin querer, así que sugieren alternativas al uso de taxonomías raciales.
La raza en la aplicación de la ley en Estados Unidos y el Reino Unido
En un intento de proporcionar descripciones generales que puedan facilitar el trabajo de la policía buscando detener a los sospechosos, el FBI emplea el término “raza” para resumir la apariencia general (color de piel, textura de pelo, forma del ojo, y otras características fácilmente observables) de los individuos que están intentando detener. Desde la perspectiva de la policía, es generalmente más importante llegar a una descripción que sugerirá fácilmente la apariencia general de un individuo que hacer una categorización científicamente válida. Así además de asignar un individuo a una categoría racial, la descripción incluirá: altura, peso, color de ojos, cicatrices y otras características distintivas, etc. Scotland Yard usa una clasificación basada en los antecedentes étnicos de la sociedad británica: W1 (británico blanco), W2 (irlandés blanco), W9 (cualquier otro origen blanco); M1 (blancos y negros del Caribe), M2 (blancos y negros africanos), M3 (blancos y asiáticos), M9 (cualquier otro origen mixto); A1 (asiático-indio), A2 (asiático-pakistaní), A3 (asiático-bengalí), A9 (cualquier otro origen asiático); B1 (negros del Caribe), B2 (negros africanos), B3 (cualquier otro origen negro); O1 (chino), O9 (cualquier otro).
En 1975, la policía británica comenzó a clasificar a los detenidos según grupos raciales que luego fueron reemplazados por un Código de Identidad:
- IC1 Blanco
- IC2 Del Mediterráneo o Hispánico
- IC3 Africano/Caribeño
- IC4 Indio/Pakistaní/Bangalí, u otras de Asia
- IC5 Chino/Japonés, o personas del sudeste asiático
- IC6 Árabe/Egipcio/Magreb
- IC0 Origen desconocido
En muchos países, está prohibido legalmente mantener datos basados en la raza, que a menudo hace cuestiones de la policía los letreros de buscados al público que incluye etiquetas como “complexión de piel oscura”, etc. Hay controversia sobre la verdadera relación entre crímenes, sus castigos asignados, y la división de la gente en las llamadas “razas”. En los Estados Unidos, la práctica de clasificar en razas ha sido dictaminado inconstitucional y también constituye una violación de los derechos humanos. Hay un activo debate respecto a la causa de una marcada correlación entre los crímenes grabados, los castigos, y la gente “racialmente dividida” del país. Muchos consideran de facto la clasificación racial un ejemplo de racismo institucional en la aplicación de la ley.
Los trabajos más recientes en la taxonomía racial basada en el análisis de clusters de ADN ha llevado a la aplicación de la ley a perseguir los sospechosos basándose en su clasificación racial derivada de su evidencia de ADN dejada en la escena del crimen. El análisis de ADN sin embargo, ha demostrado tener éxito para identificar los caracteres personales más que la supuesta raza de la víctima y de los perpetradores. En un intento de ser menos subjetivo, esta clasificación es llamada “ascendencia biogeográfica” más que “raza”, pero los términos para las categorías BGA son las mismas. La diferencia es que los marcadores informativos de la ascendencia en el ADN identifican el mestizaje de la ascendencia, no la propia identidad étnica. Por lo tanto, no pueden encajar en las “razas” de Estados Unidos. Por ejemplo, el ADN de un árabe-americano, un afrodescendiente, y un hispano de precisamente la misma mezcla genética afroeuropea sería “racialmente” indistinguible.
La raza en Francia
La legislación francesa sin embargo continúan utilizando el término de 'raza', pero prohíbe cualquier discriminación racial. En el decreto del 2 de febrero de 1990 autorizó las bases de datos según orígenes raciales de las personas aun sobre la ley de 1983 sobre los derechos y obligaciones de los funcionarios que refiere solo al origen étnico y no a la raza.
La clasificación de las razas humanas
A lo largo de la historia, desde el Antiguo Egipto hasta la actualidad se han realizado numerosos intentos de realizar clasificaciones científicas de las razas humanas. En el último siglo las aportaciones con mayor repercusión pertenecen, entre otros, a Joseph Deniker, Carleton S. Coon y especialmente H.V. Vallois en su libro Les races humaines de 1944, polémico por sus posibles implicaciones políticas en el contexto de la dominación nazi de Francia.
Véase también
- Antropología
- Antropología biológica
- Antropología física
- Clan
- El gen egoísta
- Especie
- Etnia
- Genética de poblaciones
- Grupos humanos
- Historia del concepto de raza
- Identidad cultural
- Pierre-André Taguieff
- Racismo
- Racismo científico
- Racismo oculto
- Raza
- Raza (censo de los Estados Unidos)
- Regla de una gota
- Subespecie
- Teorías raciales nazis
- Variación genética humana
Bibliografía
- Abizadeh A (2001) "Ethnicity, Race, and a Possible Humanity" World Order 33.1: 23-34.
- American Association of Physical Anthropologists (1996) AAPA statement on biological aspects of race. Am J Phys Anthropol 101:569–570
- Anonymous (1996) Style matters: ethnicity, race, and culture: guidelines for research, audit, and publication. BMJ 312:1094
- ——— (2000) Census, race, and science. Nat Genet 24:97–98
- ——— (2004) The unexamined "Caucasian". Nat Genet 36:541
- Banton M (1977) The idea of race. Westview Press, Boulder
- Bhopal R (1997) Is research into ethnicity and health racist, unsound, or important science? BMJ 314:1751–1756
- Boas 1912 "Change in Bodily Form of Descendants of Immigrants" in American Anthropologist 14: 530-562
- Bonham V, Warshauer-Baker E, Collins FS (2005) The complexity of the constructs. Am Psychol 60:9–15
- Brace 1964 "A Non-racial Approach Toward the Understanding of Human Diversity" in The Concept of Race, ed. Ashley Montagu
- Braun L (2002) Race, ethnicity, and health: can genetics explain disparities? Perspect Biol Med 45:159–174
- Brodwin P (2002) Genetics, identity, and the anthropology of essentialism. Anthropol Quart 75:323–330
- Burchard EG, Ziv E, Coyle N, Gómez SL, Tang H, Karter AJ, Mountain JL, Pérez-Stable EJ, Sheppard D, Risch N (2003) The importance of race and ethnic background in biomedical research and clinical practice. N Engl J Med 348:1170–1175
- Calafell F (2003) Classifying humans. Nat Genet 33:435–436
- Calle EE, Kaaks R (2004) Overweight, obesity and cancer: epidemiological evidence and proposed mechanisms. Nat Rev Cancer 4:579–591 * Caspari, Rachel 2003 "From Types to Populations: a Century of Race, Physical Anthropology, and the American Anthropological Association", in American Anthropologist 105(1): 65-76
- Cardon LR, Palmer LJ (2003) Population stratification and spurious allelic association. Lancet 361:598–604
- Collins-Schramm HE, et al., (2004) Mexican American ancestry-informative markers: examination of population structure and marker characteristics in European Americans, Mexican Americans, Amerindians and Asian. Human Genetics 114:263-71
- Cooper RS, Kaufman JS, Ward R (2003) Race and genomics. N Engl J Med 348:1166–1170
- Dobzhansky, T. (1970). Genetics of the Evolutionary Process. New York, NY: Columbia University Press.
- ——— (2005) Race and reification in science. Science 307:1050–1051
- Ehrlich and Holm 1964 "A Biological View of Race" in The Concept of Race, ed. Ashley Montagu.
- García Leal, A. (2013). El azar creador: la evolución de la vida compleja y de la inteligencia. Colección Metatemas núm. 126. Tusquets Editores. México. ISBN 978-607-421-468-0
- Geulen, Christian (2010) [2007]. Breve historia del racismo [Geschichte des Rassismus]. Madrid: Alianza Editorial. ISBN 978-84-206-6442-2.
- Gluckman PD, Hanson MA (2004) Living with the past: evolution, development, and patterns of disease. Science 305:1733–1736
- Goldstein DB, Chikhi L (2002) Human migrations and population structure: what we know and why it matters. Ann Rev Genomics Hum Genet 3:129–152
- Guthrie RD (1996) The mammoth steppe and the origin of mongoloids and their dispersal. In: Akazawa T, Szathmary E (eds) Prehistoric Mongoloid dispersals. Oxford University Press, New York, pp 172–186
- Hannaford I (1996) Race: the history of an idea in the West. Johns Hopkins University Press, Baltimore
- Hering Torres, Max Sebastián (2003). «’Limpieza de sangre’. ¿Racismo en la Edad Moderna?». Tiempos Modernos: Revista Electrónica de Historia Moderna 4 (9).
- Hering Torres, Max Sebastián (2011). «La limpieza de sangre. Problemas de interpretación: acercamientos históricos y metodológicos». Historia crítica (45): 32-55.
- Hooton, E.A. (1926). Methods of racial analysis. Science 63, 75–81.
- Hutchinson J, Smith AD (eds) (1996) Ethnicity. Oxford University Press, New York
- Isaac, Benajmin (2022). «Un racisme avant les races?». L’Histoire (en francés) (493): 40-42.
- Jablonski NG (2004) The evolution of human skin and skin color. Annu Rev Anthropol 33:585–623
- Keita SOY, Kittles RA (1997) The persistence of racial thinking and the myth of racial divergence. Am Anthropol 99:534–544
- Lamason RL, Mohideen MA, Mest JR, Wong AC, Norton HL, Aros MC, Jurynec MJ, Mao X, Humphreville VR, Humbert JE, Sinha S, Moore JL, Jagadeeswaran P, Zhao W, Ning G, Makalowska I, McKeigue PM, O'donnell D, Kittles R, Parra EJ, Mangini NJ, Grunwald DJ, Shriver MD, Canfield VA, Cheng KC (2005). SLC24A5, a putative cation exchanger, affects pigmentation in zebrafish and humans. Science 310: 1782-6.
- LaVeist TA (1996) Why we should continúe to study race...but do a better job: an essay on race, racism and health. Ethn Dis 6:21–29
- ——— (ed) (2002) Race, ethnicity, and health. Jossey-Bass, San Francisco
- Lee SS, Mountain J, Koenig BA (2001) The meanings of "race" in the new genomics: implications for health disparities research. Yale J Health Policy Law Ethics 1:33–75
- Lewis B (1990) Race and slavery in the Middle East. Oxford University Press, New York
- Li WH, Sadler LA (1991) Low nucleotide diversity in man. Genetics 129:513–523
- Lieberman L (2001) How "Caucasoids" got such big crania and why they shrank: from Morton to Rushton. Curr Anthropol 42:69–95
- Lieberman, Hampton, Littlefield, and Hallead 1992 "Race in Biology and Anthropology: A Study of College Texts and Professors" in Journal of Research in Science Teaching 29:301-321
- Lewontin 1973 "The Apportionment of Human Diversity" in Evolutionary Biology 6:381-397
- Lin SS, Kelsey JL (2000) Use of race and ethnicity in epidemiologic research: concepts, methodological issues, and suggestions for research. Epidemiol Rev 22:187–202
- Livingstone 1962 "On the Non-Existence of Human Races" in Current Anthropology 3: 279-281
- Lohmueller KE, Pearce CL, Pike M, Lander ES, Hirschhorn JN (2003) Meta-analysis of genetic association studies supports a contribution of common variants to susceptibility to common disease. Nat Genet 33:177–182
- Loiseau, Julien (2022). «Islam médiéval. Des peuples et des couleurs». L’Histoire (en francés) (493): 44-47.
- Long, J.C. and Kittles, R.A. (2003). Human genetic diversity and the nonexistence of biological races. Hum Biol. 75, 449–71. [8]
- Mahoney MC, Michalek AM (1998) Health status of American Indians/Alaska natives: general patterns of mortality. Fam Med 30:190–195
- Marchini J, Cardon LR, Phillips MS, Donnelly P (2004) The effects of human population structure on large genetic association studies. Nat Genet 36:512–517
- Marks J (1995) Human biodiversity: genes, race, and history. Aldine de Gruyter, New York
- Mayr, E. (1969). Principles of Systematic Zoology. New York, NY: McGraw-Hill.
- Mays VM, Ponce NA, Washington DL, Cochran SD (2003) Classification of race and ethnicity: implications for public health. Annu Rev Public Health 24:83–110
- McKeigue PM (2005) Prospects for admixture mapping of complex traits. Am J Hum Genet 76:1–7
- Meltzer M (1993) Slavery: a world history, rev ed. DaCapo Press, Cambridge, MA
- Montagu (1941). "The Concept of Race in Light of Genetics" in Journal of Heredity 23: 241-247
- Montagu (1942). Man’s Most Dangerous Myth: The Fallacy of Race
- Morales-Díaz, Enrique, and Gabriel Aquino, and Michael Sletcher, ‘Ethnicity’, in Michael Sletcher, ed., New England, (Westport, CT, 2004).
- Mörner M (1967) Race mixture in the history of Latin America. Little, Brown, Boston
- Morton NE, Collins A (1998) Tests and estimates of allelic association in complex inheritance. Proc Natl Acad Sci USA 95:11389–11393
- Mosse GL (1985) Toward the final solution, 2nd ed. University of Wisconsin Press, Madison
- Nobles M (2000) Shades of citizenship: race and the census in modern politics. Stanford University Press, Stanford
- Nordborg M (1998) On the probability of Neanderthal ancestry. Am J Hum Genet 63:1237–1240
- Nordborg M, Tavare S (2002) Linkage disequilibrium: what history has to tell us. Trends Genet 18:83–90
- Ohno S (1996) The Malthusian parameter of ascents: what prevents the exponential increase of one's ancestors? Proc Natl Acad Sci USA 93:15276–15278
- Olson S (2002) Mapping human history. Houghton Mifflin, Boston
- Oppenheimer GM (2001) Paradigm lost: race, ethnicity, and the search for a new population taxonomy. Am J Public Health 91:1049–1055
- Ossorio P, Duster T (2005) Controversies in biomedical, behavioral, and forensic sciences. Am Psychol 60:115–128
- Ota Wang V, Sue S (2005) In the eye of the storm: race and genomics in research and practice. Am Psychol 60:37–45
- Page GP, George V, Go RC, Page PZ, Allison DB (2003) "Are we there yet?": Deciding when one has demonstrated specific genetic causation in complex diseases and quantitative traits. Am J Hum Genet 73:711–719
- Parra EJ, Kittles RA, Shriver MD (2004) Implications of correlations between skin color and genetic ancestry for biomedical research. Nat Genet 36:S54–S60
- Parra EJ, Marcini A, Akey J, Martinson J, Batzer MA, Cooper R, Forrester T, Allison DB, Deka R, Ferrell RE, Shriver MD (1998) Estimating African American admixture proportions by use of population-specific alleles. Am J Hum Genet 63:1839–1851
- Parra FC, Amado RC, Lambertucci JR, Rocha J, Antunes CM, Pena SD (2003) Color and genomic ancestry in Brazilians. Proc Natl Acad Sci USA 100:177–182 [9]
- Patterson N, Hattangadi N, Lane B, Lohmueller KE, Hafler DA, Oksenberg JR, Hauser SL, Smith MW, O'Brien SJ, Altshuler D, Daly MJ, Reich D (2004) Methods for high-density admixture mapping of disease genes. Am J Hum Genet 74:979–1000
- Pfaff CL, Barnholtz-Sloan J, Wagner JK, Long JC (2004) Information on ancestry from genetic markers. Genet Epidemiol 26:305–315
- Platz EZ, Rimm EB, Willett WC, Kantoff PW, Giovannucci E (2000) Racial variation in prostate cancer incidence and in hormonal system markers among male health professionals. J Natl Cancer Inst 92:2009–2017
- Pritchard JK (2001) Are rare variants responsible for susceptibility to complex diseases? Am J Hum Genet 69:124–137
- Pritchard JK, Cox NJ (2002) The allelic architecture of human disease genes: common disease-common variant...or not? Hum Mol Genet 11:2417–2423
- Provine WB (1986) Geneticists and race. Am Zoologist 26:857–887
- Rawlings JS, Weir MR (1992) Race- and rank-specific infant mortality in a US military population. Am J Dis Child 146:313–316
- Rees JL (2003) Genetics of hair and skin color. Annu Rev Genet 37:67–90
- Reich DA, Lander ES (2001) On the allelic spectrum of human disease. Trends Genet 17:502–510
- Relethford JH (2002) Apportionment of global human genetic diversity based on craniometrics and skin color. Am J Phys Anthropol 118:393–398
- Risch N (2000) Searching for the genetic determinants in a new millennium. Nature 405:847–856
- Risch N, Burchard E, Ziv E, Tang H (2002) Categorization of humans in biomedical research: genes, race and disease. Genome Biol 3 (http://genomebiology.com/2002/3/7/comment/2007 Archivado el 24 de junio de 2006 en Wayback Machine.) (electronically published July 1, 2002; accessed August 25, 2005)
- Rivara F, Finberg L (2001) Use of the terms race and ethnicity. Arch Pediatr Adolesc Med 155:119
- Rohde D, Olson S, Chang J (2004) Modeling the recent common ancestry of all living humans. Nature 431:562–566
- Roseman CC (2004) Detecting interregionally diversifying natural selection on modern human cranial form by using matched molecular and morphometric data. Proc Natl Acad Sci USA 101:12824–12829
- Rosenberg NA, Pritchard JK, Weber JL, Cann HM, Kidd KK, Zhivotovsky LA, Feldman MW (2002) Genetic structure of human populations. Science 298:2381–2385 [10] (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
- Rotimi CN (2004) Are medical and nonmedical uses of large-scale genomic markers conflating genetics and "race"? Nat Genet 36:S43–S47
- Sallout B, Walker M (2003) The fetal origin of adult diseases. J Obstet Gynaecol 23:555–560
- Sánchez Arteaga, J. (2006). Las Teorías biológicas sobre el origen de las razas humanas (1859-1900): elementos para una crítica antropológica de la racionalidad tecnocientífica. Tesis doctoral. Madrid: Universidad Autónoma de Madrid.
- Sánchez Arteaga, Juanma (2007) La razón salvaje: tecnociencia, racismo y racionalidad. Madrid: Lengua de Trapo.
- Sarich, Vincent, and Frank Miele. Race: The Reality of Human Differences. Westview Press, 2004.
- Sartre, Maurice (2004). «Les Grecs et les Romains étaient-ils racistes?». L’Histoire (en francés) (291): 30-31.
- Schwartz M, Vissing J (2002) Paternal Inheritance of Mitochondrial DNA. N Engl J Med 347:576-580
- Shields AE, Fortun M, Hammonds EM, King PA, Lerman C, Rapp R, Sullivan PF (2005) The use of race variables in genetic studies of complex traits and the goal of reducing health disparities: a transdisciplinary perspective. Am Psychol 60:77–103
- Shipler D (1997) A country of strangers: blacks and whites in America. Knopf, New York
- Shostak S (2003) Locating gene-environment interaction: at the intersections of genetics and public health. Soc Sci Med 56:2327–2342
- Shriver, M. D. et al. (2003). Skin pigmentation, biogeographical ancestry, and admixture mapping. Hum. Genet. 112, 387–399. [11]
- Sider, Gerald 1993 Lumbee Indian Histories: Race, Ethnicity, and Indian Identity in the Southern United States
- Smedley A (1999) Race in North America: origin and evolution of a worldview, 2nd ed. Westview Press, Boulder
- Smelser N, Wilson WJ, Mitchell F (eds) (2001) America becoming: racial trends and their consequences. Vol 2. National Academy Press, Washington, DC
- Smith DJ, Lusis AJ (2002) The allelic structure of common disease. Hum Mol Genet 11:2455–2461
- Smith MW, Patterson N, Lautenberger JA, Truelove AL, McDonald GJ, Waliszewska A, Kessing BD, et al (2004) A high-density admixture map for disease gene discovery in African Americans. Am J Hum Genet 74:1001–1013
- Snowden FM (1983) Before color prejudice: the ancient view of blacks. Harvard University Press, Cambridge, MA
- Spickard PR (1992) The illogic of American racial categories. In: Root MPP (ed) Racially mixed people in America. Sage, Newbury Park, CA, pp 12–23
- Stanton W (1960) The leopard's spots: scientific attitudes toward race in America, 1815–1859. University of Chicago Press, Chicago
- Stevens J (2003) Racial meanings and scientific methods: changing policies for NIH-sponsored publications reporting human variation. J Health Polit Policy Law 28:1033–1087
- Sturm RA, Teasdale RD, Box NF (2001) Human pigmentation genes: identification, structure and consequences of polymorphic variation. Gene 277:49–62
- Takaki R (1993) A different mirror: a history of multicultural America. Little, Brown, Boston
- Tang H, Quertermous T, Rodríguez B, Kardia SL, Zhu X, Brown A, Pankow JS, Province MA, Hunt SC, Boerwinkle E, Schork NJ, Risch NJ (2005). Genetic structure, self-identified race/ethnicity, and confounding in case-control association studies. Am J Hum Genet 76, 268-75. [12]
- Tate SK, Goldstein DB (2004) Will tomorrow's medicines work for everyone? Nat Genet 36:S34–S42
- Templeton AR (1998) Human races: a genetic and evolutionary perspective. Am Anthropol 100:632–650
- Thienpont, Kristiaan and Cliquet, Robert (eds.) In-group/out-group gedrag in evolutiebiologisch perspectief, Leuven: Garant, 1999. ISBN 90-5350-970-4
- Thomas DC, Witte JS (2002) Point: population stratification: a problem for case-control studies of candidate-gene associations? Cancer Epidemiol Biomarkers Prev 11:505–512
- Tishkoff SA, Kidd KK (2004) Implications of biogeography of human populations for "race" and medicine. Nat Genet 36:S21–S27
- Todorov T (1993) On human diversity. Harvard University Press, Cambridge, MA
- Vega WA, Amaro H (1994) Latino outlook: good health, uncertain prognosis. Annu Rev Public Health 15:39–67
- Venter JC, Adams MD, Myers EW, Li PW, Mural RJ, Sutton GG, Smith HO, et al (2001) The sequence of the human genome. Science 291:1304–1351
- Wacholder S, Rothman N, Caporaso N (2002) Counterpoint: bias from population stratification is not a major threat to the validity of conclusions from epidemiological studies of common polymorphisms and cancer. Cancer Epidemiol Biomarkers Prev 11:513–520
- Wallace R, Wallace D, Wallace RG (2004) Coronary heart disease, chronic inflammation, and pathogenic social hierarchy: a biological limit to possible reductions in morbidity and mortality. J Natl Med Assoc 96:609–619
- Waters M (1990) Ethnic options: choosing identities in America. University of California Press, Berkeley
- Weatherall D (1999) From genotype to phenotype: genetics and medical practice in the new millennium. Philos Trans R Soc Lond B Biol Sci 354:1995–2010
- Whyatt RM, Rauh V, Barr DB, Camann DE, Andrews HF, Garfinkel R, Hoepner LA, Diaz D, Dietrich J, Reyes A, Tang D, Kinney PL, Perera FP (2004) Prenatal insecticide exposures and birth weight and length among an urban minority cohort. Environ Health Perspect 112:1125–1132
- Wiencke JK (2004) Impact of race/ethnicity on molecular pathways in human cancer. Nat Rev Cancer 4:79–84
- Wilson JF, Weale ME, Smith AC, Gratrix F, Fletcher B, Thomas MG, Bradman N, Goldstein DB (2001) Population genetic structure of variable drug response. Nat Genet 29:265–269
- Wilson and Brown 1953 "The Subspecies Concept and Its Taxonomic Application" in Systematic Zoology 2: 97-110
- Wolpoff M, Caspari R (1997) Race and human evolution: a fatal attraction. Simon & Schuster, New York
- Microsoft Encarta (2008)
Enlaces externos
- Does Race Exist? A proponent's perspective by George W. Gill
- The Race FAQ, Human Races and related texts by John Goodrum
- Boas's remarks on race to a general audience
- Race differences in average IQ are largely genetic, 26-Apr-2005
- Discovery of the human skin color gene SLC24A5
- US Human Genome Project on "Issues of Race"
- Is Race "Real"? - forum organized by the Social Science Research Council, includes a March 2005 op-ed article by A.M. Leroi from the New York Times advocating biological conceptions of race and responses from scholars in a variety of fields.
- Race - The power of an illusion Online companion to California Newsreel's 3-part documentary about race in society, science, and history.
- Times Online, "Gene tests prove that we are all the same under the skin", 27 October 2004.
- Catchpenny mysteries of ancient Egypt, "What race were the ancient Egyptians?", Larry Orcutt.
- Judy Skatssoon, "New twist on out-of-Africa theory", ABC Science Online, Wednesday, 14 July 2004.
- Michael J. Bamshad, Steve E. Olson "Does Race Exist?", Scientific American, December 2003
- OMB Statistical Directive 15, "Standards for Maintaining, Collecting, and Presenting Federal Data on Race and Ethnicity", Federal Register, 30 October 1997.
- Sandra Soo-Jin Lee, Joanna Mountain, and Barbara A. Koenig, "The Reification of Race in Health Research"
- Michael Root, "The Use of Race in Medicine as a Proxy for Genetic Differences"
- Richard Dawkins: Race and creation (extract from The Ancestor's Tale: A Pilgrimage to the Dawn of Life) - On race, its usage and a theory of how it evolved. (Prospect Magazine October 2004) (see also longer extract here)
- Racial & Ethnic Distribution of ABO Blood Types - bloodbook.com
- Genetic Structure, Self-Identified Race/Ethnicity, and Confounding in Case-Control Association Studies
- "Gene Study Identifies 5 Main Human Populations, Linking Them to Geography", Nicholas Wade, NYTimes, December 2002. Covering "Genetic structure of human populations", Feldman et al, Science. - "Self-reported population ancestry likely provides a suitable proxy for genetic ancestry".
- "Race as a Biological Concept" by J. Philippe Rushton
- The Race Concept: A Defense by Michael Levin
- Are White Athletes an Endangered Species? And Why is it Taboo to Talk About It? Discussion of racial differences in athletics
- Google Your Race - Google generated racial profiles.