
| Insulina | ||||
|---|---|---|---|---|

| ||||
| Estructuras disponibles | ||||
| PDB | Buscar ortólogos: | |||
| Identificadores | ||||
| Símbolo | INS (HGNC: 6081) | |||
| Identificadores externos |
||||
| Locus | Cr. 11 p15.5 | |||
| Patrón de expresión de ARNm | ||||
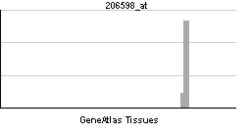
| ||||
| Más información | ||||
| Estructura/Función proteica | ||||
| Motivos |
hélice alfa lámina beta |
|||
| Ortólogos | ||||
| Especies |
|
|||
| Entrez |
|
|||
| Ensembl |
|
|||
| UniProt |
|
|||
| RefSeq (ARNm) |
|
|||
| RefSeq (proteína) NCBI |
|
|||
| Ubicación (UCSC) |
|
|||
| PubMed (Búsqueda) |
|
|||
La insulina es una hormona polipeptídica, producida y secretada por las células beta de los islotes de Langerhans del páncreas.
La insulina permite la entrada de glucosa en las células, así interviene en el control de la glucemia y en el metabolismo de los nutrientes, para la síntesis de componentes celulares como el glucógeno y los triglicéridos.
Cuando no se produce insulina suficiente o cuando el organismo del animal no utiliza eficazmente la insulina que secreta, aparecen trastornos en los niveles de la glucosa en sangre, como la #Resistencia a la insulina y la #Diabetes.
Gran número de estudios demuestran que la insulina como medicamento, es una alternativa segura, efectiva y tolerable para el tratamiento a largo plazo de la Diabetes tipo 1 y la Diabetes tipo 2.
El aislamiento y la purificación de la insulina por F. Banting en el laboratorio de Fisiología de J. Macleod fue reconocido con el Premio Nobel en 1923 para ambos.
Historia
En 1909, el investigador belga Jean de Meyer acuñó el nombre de insulina (derivado del latín insula, "isla") para designar a una sustancia, no identificada, producida por los islotes que Paul Langerhans describió, como “...montículos redondeados” como ínsulas.
En 1916 Nicolae Paulescu desarrolló en Bucarest un extracto pancreático acuoso que, tenía un efecto normalizador sobre los niveles de azúcar en la sangre. En 1921 escribió cuatro artículos sobre el trabajo realizado.
En 1921 Frederick Grant Banting, Charles Best, James Collip y J.J.R. Macleod de la Universidad de Toronto aislaron y purificaron la insulina. Otros investigadores habían producido antes extractos con efectos similares, pero se habían detenido al comprobar que causaban reacciones tóxicas. Banting y Macleod
recibieron el Premio Nobel en 1923 por el descubrimiento.
En 1923, las compañías farmacéuticas obtuvieron la licencia para producir insulina sin pagar derechos de autor, años después fue accesible.
En ese año ya se producía insulina suficiente como para comercializar. Se lograban extraer unas 3500 Unidades de insulina (U) por cada kilogramo (kg) de tejido pancreático de vaca o cerdo.
En 1951 Frederick Sanger aclaró la secuencia de aminoácidos de la insulina. Este resultado le valió su primer Premio Nobel de química en 1958.
Estructura

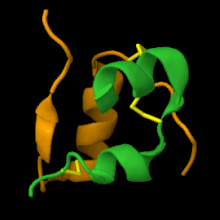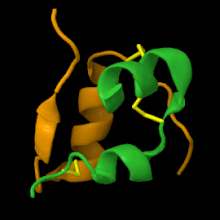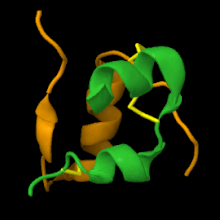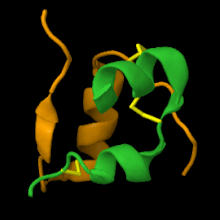
Entre los vertebrados, la insulina conserva una íntima similitud estructural. Por ejemplo, la insulina bovina difiere de la humana en solo tres aminoácidos, mientras que la porcina difiere solo en uno, por lo tanto, las insulinas de procedencia animal tienen la misma efectividad que la humana. La insulina de los túnidos es similar a la humana, y clínicamente efectiva para su uso en humanos, fue utilizada en Japón hasta 1956.
- Aún la insulina del invertebrado Caenorhabditis elegans, un nematodo, es muy similar en estructura, tiene efectos celulares muy parecidos y se produce de manera análoga a la de los humanos. De modo que es una proteína que se ha preservado a lo largo de la evolución del tiempo, sugiriendo su rol fundamental en el control metabólico animal.
- Aún la insulina del invertebrado Caenorhabditis elegans, un nematodo, es muy similar en estructura, tiene efectos celulares muy parecidos y se produce de manera análoga a la de los humanos. De modo que es una proteína que se ha preservado a lo largo de la evolución del tiempo, sugiriendo su rol fundamental en el control metabólico animal.
El péptido C, producto del desdoblamiento de la proinsulina, difiere considerablemente entre las diferentes especies.

La estructura secundaria de la insulina es muy compleja para el tamaño de la molécula, presentando motivos de estructura hélice alfa (α) y giro beta (β) en ambas cadenas y lámina beta (β) en la cadena B.
La conformación estructural de la insulina es esencial para su actividad como hormona. La insulina es sintetizada y almacenada en el cuerpo en forma de un hexámero, es decir, una unidad compuesta por seis insulinas, mientras que su forma activa es la de una hormona monómero, es decir, la molécula de insulina sola. Seis moléculas de insulina permanecen inactivas por largo tiempo en su forma hexamérica, como forma de almacenamiento de disponibilidad rápida y protección de la altamente reactiva molécula de insulina.
Genética
En el humano el gen INS, que codifica la pre-proinsulina de cadena única de 110 aminoácidos, precursora de la insulina, está localizado en el cromosoma 11 (brazo p, banda 15.5).
- Se han identificado una variedad de alelos mutantes en la región que codifica al gen. También se han descrito varias secuencias reguladoras a nivel de la región promotora del gen de la insulina humana sobre la cual se unen los factores de transcripción. En general, se sabe que las cajas A se unen a factores Pdx1, que las cajas E se unen a NeuroD, las cajas C sobre MafA y que las secuencias denominadas elementos de respuesta al cAMP se unen sobre los factores de transcripción CREB. Se han descubierto también varios silenciadores genéticos que inhiben la transcripción de la insulina.
| Secuencias reguladoras | Factores de transcripción |
|---|---|
| ILPR | Par1 |
| Caja A5 | Pdx1 |
| Elemento regulatorio negativo (NRE) | Receptor glucocorticoide, Oct1 |
| Caja Z (sobrepuesto a NRE y C2) | ISF |
| C2 | Pax4, MafA(?) |
| E2 | USF1/USF2 |
| A3 | Pdx1 |
| A2 | - |
|
CAAT enhancer binding (CEB) (parcialmente sobrepuesto a A2 y C1) |
- |
| C1 | - |
| E1 | E2A, NeuroD1, HEB |
| A1 | Pdx1 |
| G1 | - |
Síntesis
La insulina se sintetiza en las células beta (β), las cuales representan en el humano, el 60-70 % de todas las células endócrinas del páncreas.
La síntesis de la insulina se realiza a partir de su pre-pro-hormona una cadena única, que en el humano es de 110 aminoácidos y con masa molecular de 11 981 Daltons (Da), la que es transformada en pro-hormona llamada Proinsulina, y esta última en insulina.
Esta molécula precursora, por acción de enzimas proteolíticas conocidas como prohormona convertasas, específicamente la proproteína convertasa 1 y la proproteína convertasa 2, así como la exoproteasa carboxipeptidasa E.
La producción endógena de insulina es regulada en varios pasos a lo largo de su ruta sintética. Primero sobre la transcripción del ADN, específicamente a nivel del gen de la insulina. Luego a nivel de la estabilidad del ARNm y a nivel de la traducción del ARNm. Finalmente, también se regula a nivel de las modificaciones postraducción.[cita requerida]

La síntesis de la insulina pasa por una serie de etapas. Primero la pre-proinsulina es creada por un ribosoma en el retículo endoplasmático rugoso (RER), que pasa a ser (cuando pierde su secuencia señal) proinsulina. Esta es importada al aparato de Golgi, donde se modifica, eliminando una parte (péptido C) y uniendo los dos fragmentos restantes mediante puentes disulfuro.

Ciertas modificaciones ejercidas sobre la proinsulina le eliminan una región del centro de la molécula denominada péptido C quedando libres los extremos C-terminal y N-terminal. Estos extremos libres tienen 51 aminoácidos en total y se denominan cadenas A (21 aminoácidos) y B (30 aminoácidos), los cuales terminan unidas entre sí por medio de enlaces disulfuro. De modo que la proinsulina consta de las cadenas B-C-A y los gránulos secretorios liberan las tres cadenas simultáneamente.
Dentro del aparato de Golgi, la proinsulina es enviada al interior de vesículas secretoras y de almacenamiento ricas en Zn2+ y Ca2+. Una vez dentro de la vesícula, se forman especies hexaméricas de la proinsulina con dos átomos de zinc por cada hexámero de proinsulina:
(Zn2+)2(Ca2+)(Proin)6,
las cuales son posteriormente convertidas en el hexámero de insulina —por acción de enzimas proteolíticas y produciendo también la proteína C—.
(Zn2+)2(Ca2+)(In)6
La biosíntesis de insulina es generalmente rápida (menos de ∼2 h) y eficiente, con solo 1 a 2% de remanente de proinsulina dentro de los gránulos secretores maduros.

en los niveles de glucosa (en rojo).
en los niveles de insulina (en azul).
La insulina se sintetiza y se libera bajo la influencia de varios estímulos, entre ellos, la ingesta de proteínas, carbohidratos y su paso a la sangre a partir de los alimentos digeridos. Muchos carbohidratos producen glucosa, aumentando sus niveles en el plasma sanguíneo y estimulando de inmediato la liberación de insulina a la circulación portal. También se ha demostrado que la hormona de crecimiento es capaz de aumentar la secreción de insulina humana.
Se ha demostrado que la insulina y sus proteínas relacionadas son producidas también dentro del cerebro y que niveles muy reducidas de estas proteínas pueden estar asociadas a la enfermedad de Alzheimer.
Liberación de la insulina

La insulina liberada por las vesículas secretoras de las células beta (β), se vuelca directamente en el espacio intersticial, de donde pasa a los capilares fenestrados del islote y de allí a la vena porta.
La molécula activa de insulina tiene 51 aminoácidos en total y se compone de dos cadenas; una cadena A de 21 aminoácidos y una cadena B de 30 aminoácidos. Las moléculas de insulina están acompañadas en cantidad equimolecular por moléculas de péptido C.
En la circulación portal a concentraciones de ≈ 1 × 10-5 M la insulina se encuentra en forma de dímero.
En las concentraciones de aproximadamente 1 × 10-6 micromoles microM (μM) presentes en la circulación general, el monómero es la forma activa de la insulina que se une a su receptor.
Las células beta de los islotes pancreáticos liberan la hormona insulina en los diversos estados fisiológicos, ya sea en la secreción basal en ayunas o en la secreción por las ingestas de alimentos.

El páncreas en estado de ayuno (secreción basal) secreta aproximadamente una unidad por hora. Con la ingesta de alimento la secreción se incrementa de cinco a diez veces. El páncreas en su estado fisiológico secreta en total entre 40 y 50 unidades de insulina por día, unos 287 nanomoles (nmol).
Secreción basal
La secreción basal de insulina es continua, en niveles muy bajos apenas suficientes para alcanzar directamente el hígado, donde ejerce un efecto local. Esta secreción es continua entre ingestas, pero los niveles varían durante el día; los requerimientos de insulina basal son 30 % mayores en la madrugada que en las primeras horas de la noche.
Secreción aguda
La secreción de insulina provocada por las comidas, ocurre de forma rápida, es de gran magnitud y de corta duración (insulina prandial).
La primera fase de la liberación de insulina se desencadena rápidamente en respuesta al aumento de los niveles de glucosa en la sangre. La segunda fase produce una liberación sostenida y lenta de vesículas recién formadas que se activan independientemente de la cantidad de azúcar en la sangre.
En la primera fase la liberación de la insulina ocurre de manera inmediata:
- La glucosa entra en las células beta a través del transportador de glucosa GLUT2
- La glucosa pasa a la glucólisis y el ciclo respiratorio, donde se producen, por oxidación, varias moléculas de ATP de alta energía
- Los canales de potasio (K+) dependientes de los niveles de ATP y, por tanto, de los niveles de glucosa en sangre, se cierran y la membrana celular se despolariza
- Con la despolarización de la membrana, los canales de calcio (Ca2+) dependientes de voltaje se abren y el calcio entra la célula
- Un aumento en el nivel de calcio intracelular produce la activación de fosfolipasa C, que desdobla los fosfolípidos de membrana fosfatidil inositol 4,5-bifosfato en inositol 1,4,5-trifosfato y diacilglicerol
- El inositol 1,4,5-trifosfato (IP3) se une a los receptores proteicos sobre la membrana del retículo endoplásmico (RE). Esto permite la liberación de Ca2+ del RE a través de los canales IP3 aumentando más aún la concentración intracelular de calcio
- Estas cantidades significativamente mayores de calcio dentro de las células provoca la activación de la sinaptotagmina, que ayuda a la liberación de la insulina previamente sintetizada y almacenada en las vesículas secretoras.
Este es el principal mecanismo para la liberación de insulina.
La secreción de insulina es un proceso fisiológico estrechamente regulado. Esta regulación se logra por la interacción de nutrientes, hormonas gastrointestinales,
hormonas pancreáticas y neurotransmisores del sistema nervioso autónomo.
Cierta liberación de insulina ocurre además con la ingesta de otros alimentos, no solo de glucosa o hidratos de carbono, y las células beta son también influenciadas por el sistema nervioso autónomo. Los mecanismos de señalización que controlan estos vínculos son complejos.
Otras sustancias que pueden estimular la liberación de insulina incluyen los aminoácidos de las proteínas ingeridas, la acetilcolina—liberada de las terminaciones nervio vago (sistema nervioso parasimpático)—, la colecistoquinina—secretada por células enteroendocrinas de la mucosa intestinal—y el péptido insulinotrópico dependiente de glucosa (GIP). Tres aminoácidos (alanina, glicina y arginina) actúan de manera similar a la glucosa alterando el potencial de membrana de la célula beta. La acetilcolina desencadena la liberación de insulina a través de la fosfolipasa C, mientras que la colecistoquinina actúa a través del mecanismo de adenilato ciclasa.
- El sistema nervioso simpático, a través de la estimulación de receptores adrenérgicos alfa 2, como lo demuestran los agonistas de la clonidina o la alfametildopa, inhiben la liberación de insulina. Sin embargo, cabe señalar que la adrenalina circulante activará los receptores Beta 2 en las células beta de los islotes pancreáticos para promover la liberación de insulina. Esto es importante, ya que los músculos no pueden beneficiarse de los incrementos de glucosa en la sangre como consecuencia de la estimulación adrenérgica (aumento de la gluconeogénesis y glucogenolisis con los niveles bajos de la insulina en sangre: por el glucagón) a menos que la insulina está presente para permitir la translocación GLUT-4 a nivel de los tejidos. Por lo tanto, comenzando con la inervación directa, la noradrenalina inhibe la liberación de insulina a través de los receptores alfa2 y, subsecuentemente, la adrenalina circulante proveniente de la médula suprarrenal estimulará los receptores beta2-promoviendo así la liberación de insulina.
- Cuando el nivel de glucosa se reduce al valor fisiológico normal, la liberación de insulina de las células beta frena o se detiene. Si los niveles de glucosa en sangre se vuelven inferior a ese nivel, especialmente a niveles peligrosamente bajos, la liberación de hormonas hiperglicémicas, la más prominente de las cuales es el glucagón de los mismos islotes pancreáticos, pero de células alfa, obligan a la liberación de glucosa en la sangre a partir de los almacenes celulares, principalmente el almacenamiento de glucógeno en las células del hígado. Mediante el aumento de glucosa en la sangre, las hormonas hiperglucémicas previenen o corrigen la hipoglucemia que pone en peligro la vida del individuo.
La liberación de insulina está fuertemente inhibida por la hormona del estrés noradrenalina, lo que conduce a un aumento de los niveles de glucosa en sangre durante momentos de estrés.
Evidencias de alteración de la primera fase de liberación de insulina se pueden detectar en la prueba de tolerancia a la glucosa, demostrado por una sustancial elevación de nivel de glucosa en sangre en los primeros 30 minutos, un marcado descenso durante los siguientes 60 minutos, y un constante ascenso de nuevo a los niveles de referencia en las siguientes horas.
Receptor de insulina
Los receptores insulínicos (INS-R) están ubicados en diversos tipos de células del organismo, principalmente en las células del hígado, el músculo y el tejido adiposo.
Los tejidos con mayor abundancia de receptores de insulina son el parénquima hepático y el tejido adiposo, donde pueden llegar a existir 200 000 a 300 000 copias del receptor por célula.
El INS-R es un receptor transmembrana, una glucoproteína y con actividad de enzima que pertenece a los receptores de tirosina quinasa.
Este receptor está conformado por dos subunidades alfa y dos unidades beta.
En las células diana —principalmente en el hígado, músculo y tejido adiposo— se inicia una transducción de señales cuyo efecto es el incremento en la captación de glucosa y su posterior almacenamiento, evitando así un ascenso excesivo de la glucemia posprandial.
Con la reducción de la concentración circulante de glucosa, se degrada la insulina ya secretada, finalizando así la respuesta unas 2 o 3 horas después de la ingesta.
Funciones
La función de la hormona insulina es la de favorecer la entrada de glucosa desde la sangre hacia el interior de las células, principalmente: musculares (miocito), del hígado (hepatocito ) y del tejido graso (adipocito), cuando el nivel de glucosa en sangre está en ascenso.
La insulina interviene en el anabolismo, por lo que permite a las células disponer del aporte necesario de glucosa para los procesos de síntesis que implican un gasto de energía. De esta manera, mediante glucólisis y respiración celular se obtendrá la energía necesaria en forma de ATP.
- El glucagón, al contrario, es liberado a la sangre cuando el nivel de glucosa disminuye y es entonces cuando actúa.
Por su parte, la somatostatina es la hormona encargada de regular la producción y liberación tanto de glucagón como de insulina.
- Una manera de detectar si las células beta producen insulina, es una prueba para detectar la presencia del péptido C en sangre. El péptido C se libera a la sangre cuando las células beta procesan la proinsulina, convirtiéndola en insulina.
Cuando solamente entre un 10 y un 20 % de las células beta están en buen estado, comienzan a aparecer los síntomas de la diabetes, pasando primero por un estado previo denominado luna de miel, en el que el páncreas aún segrega algo de insulina.[cita requerida]
Insulina en invertebrados
En la mosca Drosophila, la vía de la insulina regula tanto el metabolismo como el crecimiento del organismo a través de un receptor de membrana único. Esta vía conjunta, que en vertebrados es la vía de la insulina y la de los péptidos parecidos a la insulina, está regulada por la ingesta de nutrientes (carbohidratos y proteínas) y por el control hormonal.
Insulina en vertebrados
En el humano la insulina tiene una importante función reguladora sobre el metabolismo, sobre el que tiene los siguientes efectos:
- Inhibe la formación de glucosa nueva (gluco-neogénesis).
- Inhibe la fragmentación del glucógeno (glucógeno-lisis) y promueve la síntesis de glucógeno (glucógeno-génesis).
- Aumenta el transporte de glucosa en el músculo esquelético y en el tejido adiposo.
- Aumenta la retención de sodio en los riñones.
- Aumenta la recaptación celular de potasio y aminoácidos.
- Disminuye la liberación de glucosa por el hígado.
- Promueve la glucólisis.
- Favorece la síntesis de triacilgleceroles (triglicéridos). Para ello, estimula la producción de acetil-CoA (por ejemplo, al acelerar la glucólisis) y también la síntesis de ácidos grasos (componentes de los triacilgliceroles) a partir de la acetil-CoA.
Los efectos fisiológicos de esta hormona son complejos y de largo alcance; en aras de la sencillez, se les ha dividido en acciones rápida, intermedia y tardía.
Acciones rápidas (segundos)
- Mayor transporte de glucosa, aminoácidos y potasio en células sensibles a la insulina.
Acciones intermedias (minutos)
- Estimulación de la síntesis de proteínas.
- Inhibición de la desintegración de proteínas.
- Activación de enzimas glucolíticas y la glucógeno sintasa.
- Inhibición de la fosforilasa y de enzimas gluconeogénicas.
Acciones tardías (horas)
- Aumento en la concentración de ARNm para las enzimas lipógenas y de otro tipo.
La fisiología de los islotes pancreáticos demuestra que la insulina por sí sola no restaura la función completa de los islotes y por qué es tan difícil lograr la normoglucemia sin la existencia de nuevos islotes que funcionen.
Patología
Resistencia a la insulina
La resistencia a la insulina es la respuesta biológica disminuida de los diferentes tejidos corporales a la hormona, con perjuicio de la efectividad de la insulina para reducir los niveles de glucosa en la sangre. Este trastorno puede conducir al desarrollo de una diabetes mellitus tipo 2.
La resistencia a la insulina se produce cuando las células de los músculos, de la grasa y del hígado no responden bien a la insulina y no pueden ingresar la glucosa de la sangre con facilidad. Entonces el páncreas se ve obligado a producir más insulina para que la glucosa siga entrando a las células (hiperinsulinemia).
- Intolerancia a la glucosa y glucemia en ayunas alterada
- La tolerancia a la glucosa (IGT) y la alteración de la glucemia en ayunas (IFG) son condiciones intermedias en la transición entre la normalidad y la diabetes.
Diabetes
La diabetes es una enfermedad crónica, que aparece cuando el páncreas no produce insulina suficiente o cuando el organismo no utiliza eficazmente la insulina que produce.
Terapias con Insulina
Insulinas animales
Luego de su descubrimiento la insulina se extrajo de páncreas de cerdo o de vaca. La insulina de estos animales es prácticamente igual a la humana, difieren en unos pocos aminoácidos. Esos extractos de páncreas contenían moléculas de proinsulina, junto con proteínas del páncreas y dímeros de la insulina.
Para evitar variaciones en la potencia del extracto y las reacciones de tipo alérgico, se creó un proceso de purificación, obteniéndose lo que se llamó insulinas monopico (o purificadas) y posteriormente, las monocomponente o altamente purificadas.
Insulinas sintéticas
Desde finales de los años 70 la insulina fue manufacturada mediante biotecnología, lográndose su síntesis a través de la recombinación del ADN recombinante con el uso de bacilos E. coli o por introducción del código genético de la pro-insulina.
En la diabetes tipo 1, y en algunos casos en la tipo 2 se hace necesaria la inyección de insulina para mantener un nivel correcto de glucosa en sangre.
Normalmente las insulinas sintéticas se sintetizan por medio de ingeniería genética a través de ADN. Hay desacuerdo sobre la eficacia de la insulina sintética comparada con la insulina derivada de las fuentes animales.
Clasificación
Existen los siguientes tipos de insulinas:
- Insulinas de acción rápida.
- Insulinas de acción corta llamada regular.
- Insulinas de acción intermedia o NPH.
- Insulinas de acción prolongada.
En muchos casos se combina el tratamiento con estos tipos de insulina.
También por su zona de inyección las podemos clasificar como:
- Insulinas subcutáneas: Cualquier insulina, ya sea de acción rápida o retardada.
- Insulinas endovenosas: Solo las insulinas de acción rápida que no poseen retardantes.
Dependiendo del retardante utilizado podemos clasificar las insulinas de la siguiente manera:
- Insulinas que utilizan zinc como retardante.
- Insulinas que utilizan otras proteínas como la protamina como retardantes.
- La conversión entre la forma hexamérica y la monomérica es una de las características fundamentales de las fórmulas de inyección de la insulina. El hexámero es mucho más estable que la hormona sola, por lo que sería una presentación más práctica, sin embargo, el monómero es la forma más reactiva de la hormona porque su difusión es mucho más rápida haciendo que no se tenga que administrar varios minutos (30-60) antes de las comidas. La presentación con la insulina más reactiva le da a los diabéticos la opción de tener comidas diarias en horas más flexibles. Ciertos preparados de insulina tienen variaciones en al menos dos aminoácidos de modo que cuando la insulina se inyecta, ésta tenga una menor tendencia de formar agregados hexaméricos y su acción sea rápida y su efecto breve.
Unidad de insulina
La unidad de insulina se define como la cantidad necesaria para disminuir la glucemia en ayunas de un conejo de 2 kg, desde 120 hasta 45 mg/dL. Se ha visto que corresponde a la 22.ª parte de un mg del producto cristalino puro adoptado como norma. O sea: 45,4545 (periodo) microgramos
Nuevos tipos de insulina
Los científicos han intentado por todos los medios conseguir tipos de insulina que no tengan que ser inyectados, procurando así hacer la vida de los enfermos algo más fácil.
Insulina inhalada
En enero de 2006 se aprobó por la Comisión Europea la primera versión de insulina inhalada para el tratamiento de la diabetes tipo 1 y tipo 2. Se trataba de la primera opción terapéutica inhalada y por tanto no inyectable desde el descubrimiento de la insulina. Se planteó como una alternativa para aquellos pacientes que por diversas razones no toleraban aceptablemente un tratamiento mediante inyecciones o pastillas.
Desde su introducción, no se consideró por algunos tan eficaz como la tradicional (subcutánea), ya que ésta se mide en centímetros cúbicos (cc) y la actual, en unidades (UI). Además al ser inhalada, no se sabe la cantidad exacta que se absorbe. Este tipo de insulina podría mejorar la calidad de vida del paciente diabético y disminuir las inyecciones y lo invasivo que resultan. No está recomendada en niños ni en ancianos. Por otra parte, no excluiría de todas las inyecciones de insulina; los diabéticos insulinodependientes deberían seguir inyectándose, siguiendo la pauta de su médico. La utilidad y valor de la insulina inhalada era más clara para quienes disfrutan de menos inyecciones en las piernas, brazos, abdomen, etc.
Sin embargo, en octubre de 2007, apenas a unos meses de haber comenzado su comercialización en España, Pfizer, laboratorio responsable de Exubera (nombre comercial de la insulina inhalada), decidió la retirada del mercado mundial del producto por no haber satisfecho sus expectativas económicas.
Noticias sobre la insulina
En 2007 se descubrió que en las células madre del cordón se producía insulina. Las células madre obtenidas del cordón umbilical de recién nacidos pueden ser manipuladas para producir insulina. La investigación fue dirigida por el Dr. Randall Urban, consiguió cultivar grandes cantidades de células madre y dirigirlas para que se asemejaran a células beta productoras de insulina, a partir de células madre adultas. El estudio según los investigadores, ofrece una alternativa al uso de células madre embrionaria.
Científicos de University of Missouri, en 2013 descubrieron que la combinación de células de la médula ósea con un nuevo medicamento, podría ayudar a curar la diabetes de tipo 1. "Hemos descubierto que la diabetes tipo 1 no sólo destruye las células productoras de insulina, sino también los vasos sanguíneos que los apoyan". El equipo de investigadores desarrolló una cura que combinaba un fármaco de creación propia con células madre adultas de la médula ósea. Parece que el fármaco detiene el ataque del sistema inmune y que las células madre generan nuevos vasos sanguíneos que ayudarían a que las células productoras de insulina se multipliquen y prosperen. En definitiva una cura de la diabetes tipo 1 necesitaría reparar los vasos sanguíneos para que las células beta crezcan y secreten la insulina.
Véase también
Bibliografía adicional
- Bliss, Michael (2013). The Discovery of Insulin (en inglés). University of Chicago Press. ISBN 9780226075631.
- Ganong, W.F. (2016). "Fisiología Médica". 25a Edición. Editorial McGraw-Hill.
Enlaces externos
- Imágenes 3D. Universidad de Salamanca
-
 Wikimedia Commons alberga una galería multimedia sobre Insulina.
Wikimedia Commons alberga una galería multimedia sobre Insulina. - En MedlinePlus hay más información sobre Insulina
| Control de autoridades |
|
|---|