
La clasificación de los virus es el proceso de nombrar virus y colocarlos en un sistema taxonómico similar a los sistemas de clasificación utilizados para los seres vivos.
Los virus se clasifican principalmente por características fenotípicas, como cápside, tipo de ácido nucleico, tipo de proteínas, ciclo replicativo, organismos huéspedes y el tipo de enfermedad que causan. Aunque las clasificaciones filogenéticas son las basadas en el tipo de proteínas, ácidos nucleicos y ciclo replicativo. La clasificación taxonómica formal de los virus es responsabilidad del sistema del Comité Internacional de Taxonomía de Virus (ICTV), aunque el sistema de clasificación de Baltimore se puede utilizar para colocar los virus en uno de los siete grupos según su forma de síntesis de ARNm. El ICTV establece convenciones de nomenclatura específicas y otras pautas de clasificación.
Estos dos métodos de clasificación no son antagónicas, y podrían integrarse totalmente entre sí, pues la clasificación del ICTV incluye algunos criterios de la clasificación de Baltimore.
Las clasificaciones en general no son monofiléticas porque los virus son un grupo polifilético. En contraposición con los seres vivos que tienen un único origen común, los virus tienen múltiples orígenes evolutivos. Según análisis filogenéticos se originaron en varias ocasiones a partir de ancestros diferentes o distintos eventos genómicos. Ver más abajo
Exclusión de los sistemas de clasificación biólogica y el árbol de la vida.
De acuerdo con la teoría celular los virus no son seres vivos, por lo que la mayoría de los científicos excluye a los virus de los sistemas de clasificación biólogica, y también se considera que por su simplicidad y rápida mutación no pueden incluirse en el árbol de la vida que describe las relaciones evolutivas entre los seres vivos. El ICTV y muchos científicos consideran a los virus como elementos genéticos móviles ya que los virus no cumplen ninguna de las funciones que definen a la vida y son más comparables a los elementos genéticos móviles que a las células (los verdaderos seres vivos). Se ha sugerido las siguientes razones para no considerar a los virus como vivos y excluirlos del árbol de la vida:
1. Los virus no están vivos. Los virus no cumplen ninguno de los criterios para cualquiera de las definiciones de vida que se han dado a lo largo de la historia desde Aristóteles hasta la actualidad. Carecen de cualquier forma de energía y metabolismo del carbono, y no pueden replicarse ni evolucionar por sí solos. Se reproducen y evolucionan únicamente dentro de las células. Sin las células del huésped, los virus son "materia orgánica compleja inanimada".2. Los virus "van por libre" filogenéticamente hablando. En un árbol filogenético, las características de los miembros de un taxón se heredan de los antepasados. Los virus no se pueden incluir en el árbol de la vida porque no comparten características con las células, y no hay un solo gen que sea compartido por todos los virus. Si bien la vida celular tiene un origen único y común, los virus son polifiléticos: tienen muchos orígenes evolutivos.
3. No hay linajes virales ancestrales. No se ha identificado un solo gen que sea compartido por todos los virus. Existen motivos proteicos comunes en las cápsides virales, pero estos probablemente se han producido a través de la evolución convergente o la transferencia horizontal de genes.
4. ¿El hecho de que los virus de hoy en día infecten a huéspedes primitivos significa que sean antiguos?. No se ha podido demostrar con certeza que los primeros virus aparecieron junto con las primeras células. El problema es que los virus se mueven fácilmente entre diversos hospedadores. La capacidad de un virus para infectar una especie en particular podría llevar a conclusiones falsas sobre la antigüedad de ese virus.
5. Los virus no tienen una estructura derivada de un ancestro común. Las células obtienen membranas de otras células durante la división celular. Por lo tanto, se puede decir que "heredan la membrana", y es correcto afirmar que las células actuales han heredado las membranas de las primeras células. Los virus no tienen esas estructuras heredadas. Todo es de nueva síntesis.
6. Los genes metabólicos virales tienen su origen en las células. Muchos genomas virales codifican proteínas involucradas en el metabolismo energético, el carbono y el metabolismo celular. Se ha argumentado que la presencia de estos genes indica que los virus son ancestros de las células. Pero no es así, los genes metabólicos no están presentes en los antepasados de estos virus. Esto implica que los virus no pueden ser ancestros de las células.
7. Los genes de traducción viral tienen su origen en las células. Algunos virus, como el caso del Mimivirus alberga genes que codifican elementos de la maquinaria de síntesis de proteínas. Se ha interpretado que esta observación significa que los virus no siempre dependieron de las células para la traducción. Sin embargo, el análisis de su secuencia indica que estos genes son derivados de las células mediante transferencia horizontal de genes, y por lo tanto no permite que los virus puedan ser incluidos en el árbol filogenético.
8. Los virus roban genes de las células. Los genomas virales codifican muchos genes que no tienen homólogos en las células. Por esto, se ha sugerido que los virus han influido en la evolución de las células al donar nuevos genes. Sin embargo, los análisis moleculares no han podido confirmar esta hipótesis. De hecho, como veremos en el siguiente punto, los virus son "ladrones de genes", no inventores de genes y proveedores genéticos masivos.
9. La transferencia genética mayoritaria es de células a virus. Como corolario a los puntos 7 y 8 podemos afirmar que el movimiento de genes es principalmente de las células a virus. La transferencia en la dirección inversa es mínima. Por lo tanto, los virus no han tenido un papel significativo en la configuración del contenido genético de las células, sino más bien al contrario, han "cogido" genes de las células y luego, mediante una capacidad de evolución y recombinación enorme, los han modificado.
10. ¿El hecho de que los virus sean simples significa que sean primitivos?. Una visión de la evolución es que es un proceso por el cual los organismos simples se vuelven más complejos. La simplicidad de muchos virus conduce a su ubicación en el origen de la vida. Sin embargo esta hipótesis ignora el hecho de que los genomas virales están sujetos a una presión selectiva para mantener un tamaño mínimo para garantizar tasas de replicación rápidas.
De todos modos, aunque la conclusión sea que los virus no están vivos y no deberían incluirse en el árbol de la vida, no implica que no sean importantes para "la vida". Los virus juegan y han jugado un papel importante en la evolución de la vida en la Tierra, aunque solo por el hecho de la presión selectiva que ejercen para que los huéspedes se defiendan y libren de ellos. Además, junto con los elementos genéticos móviles como plásmidos y transposones, pueden acabar formando parte del genoma de sus huéspedes, lo que contribuye a la creación de variabilidad genética; pueden contribuir a la transferencia horizontal de genes y por supuesto, a regular la población y la biodiversidad.
Pero, aunque sean muy importantes para la vida, su incapacidad para sostenerse y replicarse a sí mismos, su "polifiletismo", el origen celular de sus genes similares y la volatilidad de sus genomas a través del tiempo hacen imposible que incorporemos a los virus al maravilloso "Árbol de la vida".
Clasificación por morfología

Al igual que la clasificación de Baltimore basada en el genoma, los virus también se pueden clasificar basándose en su forma morfológica que puede ser icosaedra o esférica, helicoidal o filamentosa, ovoide, inusual (forma no encontrada en la mayoría de los virus) y sin cápside:
- Virus icosaedro: Ej: los adenovirus y muchos virus de Varidnaviria, herpesvirus, bacteriófagos de cola, picornavirus y relacionados, parvovirus y otros virus de Shotokuvirae, etc.
- Virus filamentoso o helicoidal: Ej: los virgavirus como el virus del mosaico del tabaco y muchos virus de ARN de plantas, los filovirus como el virus del ebola, los ronivirus y hytrosavirus de animales, los bacteriófagos del reino Loebvirae y los virus de arqueas del dominio Adnaviria.
- Virus ovoide: Ej; los polydnavirus, algunos virus gigantes como los pandoravirus y pithovirus los virus más grandes descubiertos, algunos virus de arqueas.
- Virus inusual: Ej; los rhabdovirus, los virus de la clase Naldaviricetes y muchos virus de arqueas que pueden tener formas de limón, lágrimas, espirales o globos.
- Virus sin cápside: Son virus de ARN no infecciosos y simbiontes mutualistas que se encontraron mayormente en los hongos y protistas que se clasifican en las familias Narnaviridae, Mitoviridae, Botourmiaviridae, Hypoviridae, Endornaviridae, Deltaflexiviridae, Hadakaviridae y Polymycoviridae así como el filo Taraviricota que infecta procariotas y mitocondrias eucariotas. Se consideran elementos genéticos móviles como plásmidos de ARN, pero son diferentes de ellos. La mayoría de estos virus (exceptuando a Taraviricota) se derivan evolutivamente de virus de ARN con cápsides y están compuestos de una cadena de ARN y una ARN polimerasa dependiente de ARN. Los virus sin cápsides se distinguen de los viroides y virusoides también compuestos de una cadena de ARN en que están compuestos por una ARN polimerasa dependiente de ARN y codifican proteínas en contraposición con estos que no codifican proteínas.
Clasificación de Baltimore

David Baltimore, biólogo ganador del Premio Nobel, diseñó el sistema de clasificación que lleva su nombre. El sistema de clasificación del ICTV se utiliza en combinación con el sistema de clasificación de Baltimore en la clasificación moderna de los virus.
La clasificación de Baltimore de los virus se basa en el mecanismo de producción de ARNm. Los virus deben generar ARNm de su genoma para producir proteínas y replicarse, pero cada familia de virus utiliza mecanismos diferentes. El genoma de los virus puede ser monocatenario (ss) o bicatenario (ds), de ARN o ADN, y pueden utilizar o no la transcriptasa inversa. Además, los virus ARN monocatenarios pueden ser o positivos (+) o negativos (–). Esta clasificación reparte los virus en siete grupos:
El material genético y el método por el cual los virus se replican varían entre los diferentes tipos:
Virus ADN
La replicación del genoma de la mayoría de virus ADN se produce en el núcleo de la célula. Si la célula tiene el receptor adecuado a la superficie, estos virus entran por fusión con la membrana celular o por endocitosis. La mayoría de virus ADN son completamente dependientes de la maquinaria de síntesis de ADN y ARN de la célula hospedadora y su maquinaria de procesamiento de ARN. El genoma vírico debe atravesar la envoltura nuclear de la célula para acceder a esta maquinaria.
- Virus ADN bicatenario
- Este tipo de virus tiene su material genético compuesto por ADN de doble cadena y se replica usando una ADN polimerasa, que es dependiente del ADN y no del ARN. Este tipo de virus, por lo general, debe entrar en el núcleo de la célula hospedadora antes de que sea capaz de replicarse. Además, estos virus requieren de las polimerasas de la célula hospedadora para replicar el genoma viral y, por lo tanto, son altamente dependientes del ciclo celular. Para que pueda realizarse la infección y la producción de progenie del virus se requiere que la célula esté en la fase de replicación, que es cuando las polimerasas de la célula están activas. El virus puede forzar a la célula a realizar la división celular y de forma crónica esto puede conducir a la transformación de la célula. Los virus de este grupo infectan (animales, protistas, hongos, bacterias y arqueas), sin embargo el grupo es más predominante en las bacterias y arqueas. También incluye virus satélite, virus que dependen de otros virus para su replicación. Por huéspedes, ejemplos de taxones que infectan animales son las familias; Adenoviridae, Poxviridae, Papillomaviridae, Polyomaviridae, Iridoviridae, Baculoviridae, el orden Herpesvirales, entre otras, en protistas; Mimiviridae, Phycodnaviridae, Pithoviridae, Marseilleviridae, entre otras, en hongos; Rhizidiovirus, en bacterias; Tectiviridae, Corticoviridae, Plasmaviridae, entre otras, en arqueas; Clavaviridae, Fuselloviridae, Bicaudaviridae, Globuloviridae, Ampullaviridae, Guttaviridae, el dominio Adnaviria, entre otros. Otros taxones en cambio infectan huéspedes distintos por cruzado, bacterias y arqueas; la clase Caudoviricetes y el reino Helvetiavirae, animales, hongos y protistas; Adintoviridae, animales y protistas; Asfarviridae. Ejemplos de virus satélite son los virófagos; Lavidaviridae.
- Virus ADN monocatenario
- Este tipo de virus posee en su material genético ADN de cadena sencilla y se replica usando una ADN polimerasa dependiente del ADN —al igual que el virus ADN bicatenario—. A diferencias de los virus ADN bicatenarios, estos poseen un ADN infectante monocatenario (de cadena simple), es decir, formado por una única cadena de nucleótidos, en lugar de la habitual doble hélice. Para que exista la replicación en este virus, es necesario que el ADN de cadena simple se convierta en ADN de cadena doble en las células infectadas. Los virus de este grupo infectan a todos los organismos celulares (animales, plantas, hongos, protistas, bacterias y arqueas), sin embargo son más predominantes en las bacterias y los animales. También incluye virus satélite, virus que dependen de otros virus para su replicación. Por huéspedes, ejemplos de taxones que infectan animales son las familias; Parvoviridae, Circoviridae, Anelloviridae, Bidnaviridae, Redondoviridae entre otras, en arqueas; Pleolipoviridae, Spiraviridae, en bacterias; Microviridae, Finnlakeviridae, en plantas; Nanoviridae, Geminiviridae, Metaxyviridae, entre otras, en hongos; Genomoviridae, en protistas; Bacilladnaviridae, Naryaviridae, Nenyaviridae, Vilyaviridae. Otros taxones en cambio infectan huéspedes por cruzado, bacterias y arqueas; el reino Loebvirae. Ejemplos de virus satélite son las familias Alphasatellitidae y Tolecusatellitidae.
Virus ARN
Los virus ARN son únicos porque su información genética está codificada en ARN; esto quiere decir que usan el ácido ribonucleico (ARN) como material genético, o bien que en su proceso de replicación necesita el ARN. La replicación se suele producir en el citoplasma. Los virus ARN se pueden clasificar en unos cuatro grupos según su modo de replicación. La polaridad del ARN (si puede ser utilizado directamente o no para producir proteínas) determina en gran medida el mecanismo de replicación, y si el material genético es monocatenario o bicatenario. Los virus ARN utilizan su propia ARN polimerasa dependiente de ARN para crear copias de su genoma.
- Virus ARN bicatenario
- Los virus ARN bicatenario son virus que poseen ARN de cadena doble en su genoma y no se replican usando ADN intermediario. Como la mayoría de los virus ARN, se replican en el citoplasma y no dependen de las polimerasas de las células huésped como lo hacen los virus ADN, pues incluyen estas enzimas en el virión. La traducción suele ser monocistrónica, lo que significa que cada uno de los segmentos codifica una sola proteína, a diferencia de otros virus que exhiben una traducción más compleja. Una característica particular de estos es su capacidad para llevar a cabo la transcripción de los segmentos de ARN bicatenarios bajo las condiciones apropiadas dentro de la cápside. Los virus de este grupo infectan (animales, plantas, hongos, protistas y bacterias), sin embargo el grupo es más predominante en los hongos. También incluye virus satélite, virus que dependen de otros virus para su replicación. Por huéspedes, ejemplos de taxones que infectan hongos son las familias Chrysoviridae, Quadriviridae, Hypoviridae, Megabirnaviridae, Curvulaviridae, en bacterias; Cystoviridae, Picobirnaviridae, en animales; Birnaviridae. Otros taxones en cambio infectan huéspedes distintos por cruzado, protistas y hongos; Totiviridae, Polymycoviridae, Fusariviridae, plantas y hongos; Amalgaviridae, plantas, protistas, hongos y bacterias; Partitiviridae, animales, plantas, hongos y protistas; Reoviridae, todo tipo de microorganismos; el filo Taraviricota.
- Virus ARN monocatenario positivo
- Los virus ARN monocatenarios positivos tienen ácido ribonucleico (ARN) de cadena sencilla de sentido positivo como material genético y no se replican usando ADN intermedio. Los virus ARN positivos son idénticos al ARNm y por lo tanto pueden ser inmediatamente traducidos por la célula huésped. Aunque el ARN purificado de un virus positivo puede causar directamente una infección, es menos infeccioso que el virus completo. La replicación tiene lugar principalmente en el citoplasma y no es tan dependiente del ciclo celular como en los virus ADN. Los virus ARN de sentido positivo tienen genomas con la misma polaridad del ARNm y pueden ser empleados directamente para la síntesis de proteínas usando la maquinaria de traducción de la célula huésped. Una de estas proteínas codificadas es la ARN replicasa, una ARN polimerasa que copia el ARN viral sin necesidad de pasar por una cadena de ADN intermedia. Los virus de este grupo infectan a todos los organismos celulares (animales, plantas, hongos, protistas, bacterias y arqueas), sin embargo el grupo es más predominanante en los animales y plantas. También incluye virus satélite, virus que dependen de otros virus para su replicación. Por huéspedes, ejemplos de taxones que infectan animales son las familias Picornaviridae, Caliciviridae, Flaviviridae, Astroviridae, Hepeviridae, Nodaviridae, el orden Nidovirales, entre otras, en plantas; Virgaviridae, Tombusviridae, Potyviridae, Secoviridae, Bromoviridae, Closteroviridae, entre otras, en hongos; Barnaviridae, Deltaflexiviridae, Hadakaviridae, Yadokariviridae, en protistas; Alvernaviridae, Marnaviridae. Otros taxones en cambio infectan huéspedes distintos por cruzado, bacterias y arqueas; la clase Leviviricetes, todo tipo de microorganismos; los filos Paraxenoviricota, Pomiviricota, Taraviricota y Wamoviricota, plantas y hongos; el orden Tymovirales, Botourmiaviridae, hongos y protistas; Narnaviridae y Mitoviridae, plantas, hongos y protistas; Endornaviridae. Ejemplos de virus satélite son Sarthroviridae, Albetovirus, Aumaivirus, Papanivirus y Virtovirus.
- Virus ARN monocatenario negativo
- Este virus tiene ácido ribonucleico (ARN) de cadena sencilla de sentido negativo como material genético y no se replica usando ADN intermedio. El ARN viral negativo es complementario del ARNm y por lo tanto debe convertirse en ARN positivo por una ARN polimerasa antes de la traducción. El ARN purificado de un virus negativo no es por sí mismo infeccioso, puesto que necesita ser traducido en ARN positivo. Los virus ARN de sentido negativo utilizan una ARN polimerasa o transcriptasa para formar ARN de sentido positivo. Esto significa que el virus debe aportar la enzima ARN polimerasa, puesto que esta es dependiente del ARN. La molécula ARN de sentido positivo entonces actúa como un ARNm viral, que se traduce en proteínas por los ribosomas del hospedero. Las proteína resultante se dedica directamente a la producción de los elementos de los nuevos viriones, tales como las proteínas de la cápside y la ARN replicasa, que se encarga de la producción de nuevas moléculas de ARN de sentido negativo. Los virus de este grupo infectan a todos organismos celulares (animales, plantas, hongos, protistas, bacterias y arqueas), sin embargo el grupo es más predominante en los animales y las plantas. Por huéspedes, ejemplos de taxones que infectan animales son las familias Orthomyxoviridae, Paramyxoviridae, Filoviridae, Arenaviridae, Bornaviridae, Hantaviridae, entre otras, en plantas; Aspiviridae, Fimoviridae, Tospoviridae, entre otras, en hongos; Mymonaviridae, Discoviridae, Tulasviridae, en protistas; Leishbuviridae, Yueviridae. Otros taxones en cambio infectan huéspedes distintos por cruzado, animales, plantas y hongos; Phenuiviridae, animales, plantas, hongos y protistas; Rhabdoviridae, todo tipo de microorganismos; el filo Arctoviricota.
Virus retrotranscrito
El virus retrotranscrito puede tener dos tipos de genoma:
- Virus ARN monocatenario retrotranscrito
- Un virus ARN monocatenario retrotranscrito (o virus ssRNA-RT) es un virus con ARN de cadena sencilla en su genoma que se replica en la célula hospedadora mediante transcripción inversa, es decir, mediante la formación de ADN a partir del molde ARN. Estos virus usan transcriptasa inversa codificada viralmente, es decir, una ADN polimerasa dependiente del ARN, para producir ADN a partir del genoma ARN viral. Este ADN a menudo se integra en el genoma del huésped, como en el caso de los retrovirus y pseudovirus, donde es replicado y transcrito por el huésped. Los virus de este grupo infectan a todo tipo de eucariota (animales, plantas, hongos y protistas). Por huéspedes, ejemplos de taxones que infectan un solo tipo de huésped es la familia Retroviridae que infecta animales. Las demás familias infectan huéspedes por cruzado abarcando todos los eucariotas (animales, plantas, hongos y protistas); Pseudoviridae, Metaviridae y Belpaoviridae.
- Virus ADN bicatenario retrotranscrito
- Los virus de transcripción inversa se replican mediante la transcripción inversa, que es la formación de ADN a partir de una plantilla de ARN. Los virus de transcripción inversa que contienen un genoma de ARN utilizan un intermedio de ADN para replicarse, mientras que los que contienen un genoma de ADN utilizan un intermedio de ARN durante la replicación del genoma. Este grupo contiene dos familias: Hepadnaviridae, que infecta a animales y Caulimoviridae, que infecta plantas.
Clasificación por huéspedes
Virus de células
Los virus infectan todo tipo de células y, aunque los virus existen en todo el mundo, cada especie celular tiene un grupo de virus específico, que a menudo solo infectan esta especie.
Virus de bacterias
Los bacteriófagos son un grupo extremadamente común y diverso de virus. Por ejemplo, los bacteriófagos son la forma más común de entidad biológica en los medios acuáticos; en los océanos hay hasta diez veces más de estos virus que de bacterias, alcanzando niveles de 250 millones de bacteriófagos por milímetro cúbico de agua marina. Estos virus infectan bacterias específicas uniéndose a moléculas receptoras de superficie y entrando en la célula. En un periodo corto de tiempo (en algunos casos en unos minutos), las polimerasas bacterianas empiezan a traducir ARN vírico en proteína. Estas proteínas se convierten en nuevos viriones dentro de la célula, proteínas colaboradoras que ayudan a parecerse nuevos viriones, o proteínas implicadas en la lisis celular. Las enzimas víricas colaboran en la destrucción de la membrana celular y, en el caso del fago T4, un poco más de veinte minutos después de la inyección ya se pueden liberar más de 300 fagos.
El mecanismo principal por el que las bacterias se defienden de los bacteriófagos es produciendo enzimas que destruyen el ADN ajeno. Estas enzimas, llamadas endonucleases de restricción, fragmentan el ADN vírico que los bacteriófagos introducen en las células bacterianas. Las bacterias también tienen un sistema que utiliza secuencias CRISPR para retener fragmentos del genoma de virus que se han encontrado en el pasado, lo que les permite impedir la replicación del virus mediante una forma de interferencia del ARN. Este sistema genético proporciona a las bacterias una inmunidad adquirida a las infecciones.
Virus de arqueas
Al igual que los bacteriófagos infectan bacterias, algunos virus se replican dentro de las arqueas. Esto incluye virus con formas ya conocidas en las bacterias y eucariotas y otros con una variedad de formas inusuales, como botellas, barras con un gancho o incluso lágrimas que no parecen tener relación evolutiva con otros virus, estos últimos han sido descritos en mayor detalle en los termófilos, en particular los órdenes Sulfolobales y Thermoproteales. La defensa contra estos virus pueden incluir la interferencia del ARN de secuencias de secuencias repetidas de ADN del genoma arqueobacterial que están relacionadas con los genes de los virus.
Virus de protistas
Los virus protistas pueden infectar a diversos organismos eucariotas primitivos como algas, protozoos, mohos mucilaginosos y pseudohongos. Se desconoce mucho las características de los virus protistas, pero pueden transmitirse por esporas, difusión pasiva, reproducción y fagocitosis de partículas virales. Los virus protistas pueden jugar un papel importante en el control de la proliferación de algas y la población de protozoos, incluyendo protozoos parásitos. Los protistas se destacan por albergar los virus más grandes conocidos que infectan principalmente amebas y mohos mucilaginosos.
Virus de hongos
Los efectos de las infecciones por micovirus pueden variar de ventajosas a perjudiciales, pero la mayoría de los micovirus son asintomáticos o crípticos. En consecuencia, los síntomas solo pueden aparecer cuando ciertas condiciones del sistema virus-hongo cambian y se desequilibran. Esto podría ser externo (ambiental) así como interno (citoplasmático). Todavía no se sabe por qué algunas combinaciones de micovirus y hongos suelen ser perjudiciales, mientras que otras son asintomáticas o incluso beneficiosas. Se desconoce si el transporte viral es un proceso activo o pasivo, generalmente se supone que los virus fúngicos avanzan por flujo de plasma. Es posible que los micovirus simplemente no necesiten una ruta externa de infección, ya que tienen muchos medios de transmisión y propagación debido al estilo de vida de su huésped fúngico como la plasmogamia e intercambio citoplasmático durante largos períodos de tiempo, producción de grandes cantidades de esporas asexuales, pasar el invierno a través de esclerocios y transmisión más o menos efectiva en esporas sexuales.
Virus de plantas
Hay muchos tipos de virus de las plantas o fitovirus, pero a menudo solo causan una pérdida de producción, y no es económicamente viable intentar controlarlos. Los virus de las plantas a menudo son transmitidos de una planta a otra por organismos conocidos como vectores. Normalmente son insectos, pero también se ha demostrado que algunos hongos, nemátodos y organismos unicelulares son vectores. Cuando se considera económico el control de infecciones por fitovirus (en los frutos perennes, por ejemplo), los esfuerzos se concentran en matar a los vectores y eliminar huéspedes alternativos como malas hierbas.
Las plantas tienen mecanismos de defensa elaborados y eficientes contra los virus. Uno de los más eficientes es la presencia de los llamados genes de resistencia (R). Cada gen R confiere resistencia a un virus determinado desencadenando zonas localizadas de muerte celular alrededor de la célula infectada, que se pueden ver a simple vista en forma de manchas grandes. Esto detiene la expansión de la infección. La interferencia del ARN también es una defensa efectiva en las plantas. Cuando están infectadas, las plantas a menudo producen desinfectantes naturales que matan los virus, como el ácido salicílico, el óxido nítrico y moléculas reactivas de oxígeno.
Virus de animales
Los virus son importantes patógenos del ganado. Enfermedades como la fiebre aftosa y la lengua azul son causadas por virus. Los animales de compañía (como perros, gatos y caballos), si no se les vacuna, son susceptibles a infecciones víricas graves. El parvovirus canino es causado por un pequeño virus ADN y las infecciones a menudo son fatales en los cachorros. Como todos los invertebrados, la abeja de la miel es susceptible a muchas infecciones víricas. Afortunadamente, la mayoría de virus coexisten de manera inofensiva con su huésped y no causan signos o síntomas de enfermedad.
Virus con más de dos tipos de huéspedes distintos
Algunos virus pueden infectar a dos tipos de huéspedes distintos por cruzado como Reoviridae y Rhabdoviridae; animales, plantas, hongos y protistas pasando por el ser humano y organismos unicelulares como los protozoos. Muchos virus asociados a huéspedes distintos son virus de hongos que a la vez se encuentran asociados con plantas como Alphaflexiviridae, Betaflexiviridae, Tymoviridae y Amalgaviridae, o protistas como Totiviridae, Narnaviridae, Mitoviridae y Fusariviridae. Los caudovirus (Caudoviricetes) o los famosos bacteriófagos de cola no solo infectan a las bacterias, sino también a arqueas y por lo tanto infectan huéspedes distintos por cruzado que serían células procariotas en contraposición con los anteriores que infectarían células eucariotas.
Virus satélite
A pesar de que los virus sean dependientes de las células para su supervivencia, existen virus que dependen de otros virus para su existencia, denominados virus satélite que no pueden replicarse sin ayuda de un virus auxiliar y son más pequeños que los virus comunes. Los virus satélite generalmente necesitan un virus auxiliar del cual requeriran en parte la maquinaria replicativa (polimerasas y otras enzimas) del virus huésped, generalmente no son dañinos para sus virus auxiliares. Sin embargo algunos como los virófagos si pueden ser parásitos o infectivos al inhibir su replicación, secuestrar totalmente su maquinaria replicativa, causarles malformación en su desarrollo y suprimir su número de viriones. Aunque los virus satélite se repliquen en un huésped viral, no pueden comenzar a hacerlo hasta que su virus auxiliar haya infectado a la célula huésped. Se cree que se podrían insertar en los genomas virales a modo de transposones víricos.
Los virus satélite también pueden defender a las células huéspedes de sus virus, por ejemplo los alfasatélites y tolecusatélites pueden controlar a sus virus auxiliares reduciéndoles la virulencia y la infectividad cuando tomen su maquinaria replicativa. Por otra parte, los virófagos se replican secuestrando los aparatos de replicación de los virus auxiliares, suprimiendo así el número de viriones producidos por los virus auxiliares, aumentando la probabilidad de supervivencia del huésped. Algunos virófagos pueden endogenizarse, y esta endogeneización puede considerarse una forma de inmunidad adaptativa para el huésped contra la infección por sus virus.
Agentes subvirales


Se denomina agente subviral a un agente biológico de tipo acelular considerado similar a un virus, pero que no cumple con todas las características típicas de los virus, por lo que se presume que son más simples que aquellos.
Generalmente se considera que un virus típico es un organismo submicroscópico, infeccioso, intracelular y que está constituido por ácidos nucleicos (ADN o ARN) envueltos en una cubierta glicoproteica llamada cápside. En este sentido, un agente subviral se define negativamente como aquella partícula que no cumple completamente con la definición de un virus típico, por lo que los agentes virales serían los siguientes:
- Satélite: Virus que depende o parasita a otro virus a manera de coinfección dentro de una célula huésped. Un ejemplo es el grupo de los virófagos.
- Viroide: Virus que no presenta cápside, por lo que el ácido nucleico está desnudo.
- Virusoide: Al igual que el viroide, el ácido nucleico está desnudo, y como el satélite, necesita parasitar otro virus.
- Elemento viral endógeno: Secuencia genética viral integrada a una célula. Un ejemplo son los provirus.
- Partícula defectiva interferente: Porción de ARN o ADN viral generado por mutación que ocurre durante una replicación defectuosa y que disminuye el potencial infeccioso viral.
- Partícula similar a virus: Partícula proteica de un virus, no infecciosa y sin ácido nucleico, para su uso en vacunas.
- Prion: Agente proteico autorreplicante infeccioso cuando su estructura secundaria está modificada.
De estos agentes subvirales los viroides, satélites y virusoides son reconocidos en la taxonomía del ICTV. Los priones también están reconocidos por el ICTV pero se les asigna una clasificación no taxonómica aparte.
Clasificación del ICTV
Esquema y pautas

El Comité Internacional de Taxonomía de Virus comenzó a diseñar e implementar reglas para el nombre y la clasificación de virus a principios de la década de 1970, un esfuerzo que continúa hasta el presente. El ICTV es el único organismo encargado por la Unión Internacional de Sociedades Microbiológicas con la tarea de desarrollar, refinar y mantener una taxonomía de virus universal. El sistema comparte muchas características con el sistema de clasificación de organismos celulares como la estructura del taxón. Sin embargo, existen algunas diferencias, como el uso universal de las cursivas para todos los nombres taxonómicos, a diferencia del Código Internacional de Nomenclatura para algas, hongos y plantas, el Código Internacional de Nomenclatura de Bacterias y el Código Internacional de Nomenclatura Zoológica.
La clasificación viral comienza en el nivel de dominio y continúa de la siguiente manera, con los sufijos taxonómicos entre paréntesis.
La estructura general de la taxonomía es la siguiente:
- Dominio (-viria)
- Reino (-virae)
- Filo (-viricota)
- Subfilo (-viricotina)
- Clase (-viricetes)
- Orden (-virales)
- Suborden (-virineae)
- Familia (-viridae)
- Subfamilia (-virinae)
- Género (-virus)
- Especie (-virus)
A diferencia del sistema de nomenclatura binomial adoptado en las especies de seres vivos, actualmente no existe una forma estandarizada para los nombres de las especies de virus. En la actualidad, la ICTV exige que el nombre de una especie debe contener la menor cantidad de palabras posible sin dejar de ser distinto, y no solo debe contener la palabra virus y el nombre del huésped. Los nombres de las especies a menudo toman la forma de virus enfermedad, particularmente para los virus de plantas y animales superiores. En 2019, la ICTV publicó una propuesta para adoptar un sistema más formalizado de nomenclatura binomial para los nombres de las especies de virus, que se votó en 2020. Sin embargo, algunos virólogos luego objetaron el posible cambio del sistema de nombres, argumentando que el debate se produjo mientras que muchos en el campo estaban preocupados debido a la pandemia de COVID-19.
Definición de especies virales
Las especies forman la base de cualquier sistema de clasificación biológica. Antes de 1982, se pensaba que los virus no podían adaptarse al concepto reproductivo de especie de Ernst Mayr, por lo que no eran susceptibles de tal tratamiento. En 1982, la ICTV comenzó a definir una especie como "un grupo de cepas" con cualidades de identificación únicas. En 1991, se adoptó el principio más específico de que una especie de virus es una clase politética de virus que constituye un linaje en replicación y ocupa un nicho ecológico particular.
En julio de 2013, la definición de especie de ICTV cambió para declarar: "Una especie es un grupo monofilético de virus cuyas propiedades pueden distinguirse de las de otras especies por múltiples criterios". Los virus son entidades físicas reales producidas por la evolución biológica y la genética, mientras que las especies de virus y los taxones superiores son conceptos abstractos producidos por el pensamiento racional y la lógica. Por tanto, la relación virus / especie representa la primera línea de la interfaz entre biología y lógica.
Los criterios reales utilizados varían según el taxón y pueden ser inconsistentes (umbrales de similitud arbitrarios) o no relacionados con el linaje (geografía) en ocasiones. Para muchos, el asunto aún no está resuelto.
Definición del ICTV para cada entidad viral
Actualmente el ICTV reconoce en su taxonomía, aparte de los virus a los viroides, virusoides y satélites. Propone la siguiente definición para cada entidad viral y su clasificación:
El ICTV define virus y su clasificación de la siguiente manera:
Los virus se definen operacionalmente por el ICTV como un tipo de elemento genético móvil que codifica al menos una proteína que es un componente principal del virión que encierra el ácido nucleico del elemento genético móvil respectivo y, por lo tanto, el gen que codifica la proteína del virión principal en sí; o elemento genético móvil que se pueda demostrar claramente que son miembros de una línea de ascendencia evolutiva de las principales entidades que codifican la proteína del virión. Cualquier grupo monofilético de elementos genéticos móviles que se origine a partir de un antepasado que codifica la proteína del virión debe clasificarse como un grupo de virus.
El ICTV define viroide y su clasificación de la siguiente manera:
Los viroides se definen operacionalmente por el ICTV como un tipo de elemento genético móvil de ARN monocatenario, circular, pequeño y sin recubrimiento que no codifica proteínas y no dependen de virus para la transmisión, y que se replican de forma autónoma a través de un mecanismo en círculo rodante de ARN-ARN mediado por enzimas del huésped y, en algunos casos, por ribozimas con cabeza de martillo propias que actúan en cis; o elemento genético móvil que se derivan de un viroide en el curso de la evolución. Cualquier grupo monofilético de elemento genético móvil que se origine a partir de un antepasado viroide debe clasificarse como un grupo de viroides.
El ICTV define satélite/virusoide y su clasificación de la siguiente manera:
Los satélites/virusoides se definen operacionalmente por el ICTV como un tipo de elemento genético móvil no viroides, que dependen de los virus para su replicación y transmisión; o elemento genético móvil que se derivan de tales entidades en el curso de la evolución. Cualquier grupo monofilético de elemento genético móvil que se origine a partir de un antepasado satélite/virusoide debe clasificarse como un grupo de satélite/virusoide.
Los virus, viroides y virusoides/satélites tienen una historia evolutiva aliada a los elementos genéticos móviles no virales por lo que pudieron dar origen a algunos de ellos, pero también los elementos genéticos móviles no virales pudieron originar a ciertas entidades virales. Algunos elementos genéticos móviles no virales derivados de virus están incluidos en la taxonomía como los retrotransposones LTR incluidos en las familias Pseudoviridae, Metaviridae y Belpaoviridae derivados de los virus retrotranscritos, los virus sin cápsides o plásmidos de ARN derivados de los virus de ARN; Narnaviridae, Mitoviridae, Botourmiaviridae, Hypoviridae, Endornaviridae, Deltaflexiviridae, Polymycoviridae y Hadakaviridae, los agentes de transferencia de genes incluidos en las familias Bartogtaviriformidae, Brachygtaviriformidae y Rhodogtaviriformidae. También se ha propuesto incluir a los transposones polintones derivados de los virus de Varidnaviria, los helitrones derivados de Monodnaviria, los retrotransposones retrozimas derivados de los viroides así como otros grupos de elementos genéticos móviles descendientes de las entidades virales.
Historia evolutiva y dominios
La taxonomía actual del ICTV está basada en el tipo de proteínas que posean y la secuencia de aminoácidos. Según esta clasificación los virus pueden dividirse en 6 dominios y algunos reinos. Las hipótesis más aceptadas por el ICTV para el origen de los virus son la del escape y el virus primero hechas un híbrido, ya que estas dos hipótesis explican su simplicidad, sus características, sus genomas y genes diferentes a veces asociados con elementos genéticos móviles y genomas celulares, en algunas ocasiones sin homólogos, sus proteínas y enzimas variadas algunas asociadas con las células y la mayoría sin homólogos, sus membranas lípidicas derivadas de las celulares, su módulo de replicación y estrategias replicativas ausentes en las células, los genomas variados (ver clasificación de Baltimore).
A continuación se especifican los dominios virales de acuerdo con sus características y evolución, según lo expuesto por sus promotores.
Dominios de ARN

-
Riboviria: Contiene los virus que codifican ARN polimerasas dependientes de ARN y transcriptasas inversas. La agrupación surge porque se cree que las ARN polimerasas dependientes de ARN y transcriptasas inversas comparten un ancestro común. Durante el antiguo mundo de ARN estas enzimas codificadas por ciertos replicones precederían al ADN incluyendo los virus de ADN. Incluye los virus de ARN y los virus retrotranscritos que se clasifican en sus propios reinos aunque algunas familias y géneros de virus de ARN no están asignados.
- Orthornavirae: Incluye los virus de ARN con excepción de algunas familias y géneros poco investigados. Codifican una ARN polimerasa dependiente de ARN lo que permite estudiar toda su filogenia y evolución. Esta enzima se caracteriza por replicar ARN-ARN sin transcripción de ADN (una característica ausente en las células). El ancestro de los virus de ARN fue un replicón precelular de ARN similar a los mitovirus y endornavirus (sin cápside) que codificaba una RdRP. Este replicón al fusionarse con la RdRP y proteínas de dominio palma, TNF, Pro-PD, CBM, nucleoplasminas y una JRC-MCP horizontal llevó a la formación de los viriones de Orthornavirae. Se ha sugerido que los virus de ARN precedieron a los intrones y retrotransposones en el mundo precelular mediante un ancestro compartido con el filo Lenarviricota. Según estos estudios el filo más divergente de los virus de ARN es Taraviricota descubierto por metagenómica los cuales serían virus de ARN sin cápside y que infectarían procariotas y mitocondrias eucariotas. Los eucariovirus de ARN descienden de los arqueovirus y bacteriovirus de ARN donde tuvieron una gran diversificación que representan la gran cantidad de virus que infectan eucariotas. Sus miembros pueden tener cápsides icosaedricas o helicoidales con envoltura o sin envoltura.
- Pararnavirae: Incluye los virus retrotranscritos. Codifican una transcriptasa inversa que permite estudiar toda su filogenia y evolución. Se originaron de un evento en el que un retrotransposón se integró en la cápside de un virus de ARN, remplazando el genoma y las enzimas típico del virus. Sus virus tienen cápsides icosaédricas en ciertos casos con envoltura, sin embargo algunos de sus miembros como los retrotransposones LTR son retrotransposones derivados de estos que codifican proteínas virales.
- Ribozyviria: Incluye a los virusoides, viroides de tipo satélite que dependen de virus de ARN y retrotranscritos, principalmente de animales, plantas, bacterias y hongos, para su replicación. Se caracterizan por ser moléculas de ARN que se replican a través de ribozimas, aunque en algunos miembros una parte de su ciclo implica utilizar la polimerasa propia del virus huésped (RdRP o RT), algunos de ellos obtienen un ensamblaje viral a partir del que porta el virus huésped. Incluye al agente causante de la Hepatitis D. Los análisis filogenéticos y de homología basados en diferentes tipos de ARNs sugieren que los virusoides comparten un origen monofilético con los viroides y que ambos descienden de un ancestro común. Los análisis filogenéticos también sugieren que las entidades de tipo viroide dieron origen a las retrozimas, un tipo de retrotransposón que se replica a través de una ribozima con cabeza de martillo presente en las plantas y animales y a los retroviroides un tipo de elemento genético móvil derivado de un viroide unido con el ADN y transcriptasa inversa de un caulimovirus, encontrado en los claveles. Los virusoides y viroides se consideran reliquias de la primera fase del mundo de ARN, porque ambos carecen de homología con moléculas de ARN celulares, su replicación se basa únicamente en el ARN (sin pasar por un extremo o secuencia de ADN, una característica ausente en las células), no codifican proteínas y tienen propiedades químicas propuestas para el mundo de ARN y el caldo primigenio y ambos se replican en su mayor parte a través de ribozimas consideradas las primeras moléculas de ARN en surgir.
- Viroidia: Incluye a los viroides que son agentes infecciosos compuestos por una molécula de ARN sin cápside o polimerasas que no codifican proteínas y no transcriben ADN. Se replican mediante un mecanismo de círculo ARN-ARN mediado por enzimas del huésped o ribozimas propias. Los viroides infectan plantas y recientemente se han reportado infectando hongos, sin embargo también han demostrado poder replicarse en bacterias y algas. En 2022 un estudio de muestras metagenómicas de diversos hábitats tanto acuáticos como terrestres ha detectado muchos ARN viroides no descubiertos que infectarían otros huéspedes que no son plantas como hongos, eucariotas unicelulares (algas unicelulares y protozoos) y procariotas (bacterias y arqueas). Los análisis filogenéticos y de homología basados en diferentes tipos de ARNs sugieren que los viroides comparten un origen monofilético con los virusoides y que ambos descienden de un ancestro común. Los análisis filogenéticos también sugieren que las entidades de tipo viroide dieron origen a las retrozimas, un tipo de retrotransposón que se replica a través de una ribozima con cabeza de martillo presente en las plantas y animales y a los retroviroides un tipo de elemento genético móvil derivado de un viroide unido con el ADN y transcriptasa inversa de un caulimovirus, encontrado en los claveles. Los viroides junto con los virusoides se consideran reliquias de la primera fase del mundo de ARN, porque ambos carecen de homología con moléculas de ARN celulares, su replicación se basa únicamente en el ARN (sin pasar por un extremo o secuencia de ADN, una característica ausente en las células), no codifican proteínas, tienen propiedades químicas propuestas para el mundo de ARN y el caldo primigenio y ambos se replican en su mayor parte a través de ribozimas consideradas las primeras moléculas de ARN en surgir.
Dominios de ADN

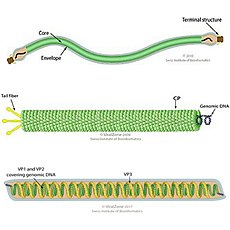


- Duplodnaviria: Incluye virus de ADN que tiene una proteína específica denominada HK97-MCP y una enzima única la terminasa. Sus virus son de ADN bicatenario y con cápsides icosaedricas. Contiene los clásicos caudovirus procariotas y sus descendientes eucariotas los herpesvirus y los mirusvirus. Se ha sugerido que se originaron antes que el último antepasado común universal (LUCA). La fusión de un replicón de ADN precelular con una ADN polimerasa, la terminasa, la HK97-MCP y proteínas de dominio DUF1884, hélice-hebra, dodecinas y encapsulinas llevó a la formación de los viriones de Duplodnaviria, que al principio no tendrían la cola contráctil, por lo que esta se habría desarrollado después de LUCA. Los herpesvirus y mirusvirus que infectan eucariotas descienden de los caudovirus con la pérdida de la cola constractil.
- Adnaviria: Incluye virus de arqueas que contienen una proteína específica dímera denominada SIRV2 dentro la cápside, morfología filamentosa y ADN en forma de A. Todos los representantes son virus de ADN bicatenario. Se ha sugerido que se originaron antes que el último antepasado común universal (LUCA). La fusión de un replicón de ADN precelular con la ADN polimerasa, la SIRV2 y proteínas de dominio hélice-hélice, METT26, SER2, ILE26 y ARG128 llevó a la formación de los viriones de Adnaviria. Es también el único dominio que hasta ahora no tiene descendientes en los eucariotas.
-
Varidnaviria: Incluye virus de ADN que tienen una proteína específica en rollo de gelatina vertical. Pueden ser en doble rollo de gelatina vertical DJR-MCP o en rollo simple de gelatina SJR-MCP que conforman sus propios reinos. Sus miembros son virus de ADN bicatenario que contiene cápsides mayormente icosaedricas. Se ha sugerido que se originaron antes que el último antepasado común universal (LUCA). La fusión de un replicón de ADN precelular con la ADN polimerasa y proteínas de dominio cupin, TNF, Pro-PD, CBM, nucleoplasminas y la SJR-MCP vertical llevó a la formación de los viriones de Varidnaviria. Los eucariovirus descienden de tectivirus a través de unos virus intermediarios llamados "polintovirus" que posteriormente se convertirían en los transposones polintones.
- Bamfordvirae: Incluye los virus con la proteína en doble rollo de gelatina DJR-MCP que derivan de los virus con proteínas en rollo simple de gelatina SJR-MCP. Sus virus tienen principalmente cápsides icosaédricas, aunque en algunos virus gigantes y la clase Naldaviricetes la forma ancestral fue remplazada por formas inusuales como ovoides o bastones. Contiene varios grupos de virus procariotas junto con todos los eucariovirus del dominio como los adenovirus, los virus gigantes y los virofagos. Los eucariovirus descienden de los bacteriófagos tectivirus a través de unos virus intermediaros denominados "polintovirus" que posteriormente se convertirían en los transposones polintones. Los virus gigantes evolucionaron de virus pequeños descendientes de los polintovirus, que aumentaron su genoma y el tamaño del virión mediante la duplicación y deleción de genes, la inclusión de elementos genéticos móviles y la adquisición masiva de genes del huésped, incluidos los genes para la traducción y los genes informáticos que se consideran los más resistentes a la transferencia horizontal. Los virus gigantes pudieron haber dado origen a los virus inusuales de la clase Naldaviricetes o descender de un ancestro compartido.
- Helvetiavirae: Incluye los virus con la proteína en rollo de gelatina simple SJR-MCP que fueron los primeros virus de Varidnaviria y que precedieron a los virus DJR-MCP. Mantienen la cápside icosaédrica ancestral y todos ellos son virus procariotas.
-
Monodnaviria: Incluye virus de ADN que tienen una proteína específica denominada rep que hace una replicación en círculo rodante y una endonucleasa HUH. Casi todos sus miembros son virus de ADN monocatenario. Se originaron de tres eventos en los que plásmidos procariotas se integraron en las cápsides de virus de ARN monocatenario positivo del filo Kitrinoviricota, remplazando el genoma y las enzimas típico del virus.
- Shotokuvirae: Contiene los eucariovirus del dominio ejemplo los papilomavirus, poliomavirus, parvovirus, circovirus, nanovirus, geminivirus, bacilladnavirus etc. Se originaron de bacteriófagos similares a los microvirus del reino Sangervirae. Sus virus son todos con cápsides icosaedricas sin envoltura vírica y con genomas circulares. Una de sus familias los bidnavirus se originaron de un evento en el que las proteínas de la cápside de los parvovirus se integraron en el genoma de un transposón polinton remplazando varias características típicas de Monodnaviria.
- Sangervirae: Contiene bacteriófagos con cápsides icosaédricas sin envoltura vírica y genomas circulares. Se originaron de ciertos plásmidos procariotas independientemente de los otros reinos y son el origen de los eucariovirus del dominio.
- Trapavirae: Contiene virus de arqueas con cápsides icosaédricas cubiertas de una envoltura vírica y genomas circulares. Se originaron de ciertos plásmidos procariotas independientemente de los otros reinos.
- Loebvirae: Contiene virus procariotas con cápsides helicoidales y genomas circulares. Se originaron de ciertos plásmidos procariotas independientemente de los otros reinos.
- Linaje de virus fusiformes y Clavaviridae: Se ha sugerido que las familias de virus fusiformes o en forma de limón de arqueas Fuselloviridae, Halspiviridae, Thaspiviridae, Itzamnaviridae y Bicaudaviridae están relacionadas por la presencia de una proteína homóloga la SSV1, ATPasas de la superfamilia AAA, forma de limón y el ensamblaje, la proteína homóloga también está presente en la familia de virus arqueanos filamentosos Clavaviridae, por lo que se ha sugerido que estos virus evolucionaron de virus filamentosos emparentados con Clavaviridae que alteraron su forma para almacenar un genoma más grande y conformarían un dominio o linaje. Su origen al igual que otros dominios es anterior al último antepasado común universal (LUCA).
Además existen 16 familias de virus de ADN de arqueas que no han sido asignadas a ningún dominio debido a que no tienen proteínas o genes homológos con otros virus o los elementos genéticos móviles no virales, por tanto se consideran que están desconectados de la virosfera, aunque el escenario más probable es que su desconexión se deba a que son descendientes de linajes de virus primordiales extintos que se habrían originado antes que el último antepasado común universal (LUCA), al igual que los otros dominios. Estos virus son en su gran mayoría virus de ADN bicatenario que tienen como característica cápsides con formas inusuales como ovoides, gusanos, flautas, botellas, lagrimas, pipas o helicoides raras que contrastan con las cápsides de otros virus. Algunos virus de arqueas también se han podido relacionar entre sí como el dominio Adnaviria que incluye los virus filamentosos de arqueas. Algunos virus inusuales de arqueas como Ampullaviridae, Bicaudaviridae, Guttaviridae y Fuselloviridae, Thaspiviridae y Halspiviridae parecen estar relacionados con plásmidos y transposones procariotas y de hecho sus genomas pueden haber evolucionado a partir de estos elementos genéticos móviles no virales. Las familias Ampullaviridae, Thaspiviridae y Halspiviridae muestran una relación estrecha con los transposones capsones, mientras que las familias Guttaviridae, Bicaudaviridae, Fuselloviridae, están vinculadas con algunos tipos de plásmidos arqueales. Se ha planteado la posibilidad de que la enigmatica virosfera arqueal evoluciono mediante la recombinación genética entre elementos genéticos móviles no virales y virus. También queda sin asignar la familia de virus bacterianos de ADN inusuales Plasmaviridae que tampoco muestra relación filogenética con otros virus y organismos celulares, cuya procedencia podría ser similar a la que se propuso para los virus inusuales de arqueas. Por último también queda por rellenar la clasificación taxonómica y el origen de varios virus satélite y los viroides vacío que podrá completarse a largo plazo gracias a la investigación moderna.
Taxonomía
La taxonomía actualizada por el Comité Internacional de Taxonomía de Virus en 2023 es la siguiente:
- Dominio Riboviria
- Reino Pararnavirae
- Familia Hepadnaviridae
- Orden Ortervirales
- Familia Belpaoviridae
- Familia Caulimoviridae
- Familia Retroviridae
- Familia Metaviridae
- Familia Pseudoviridae
- Reino Orthornavirae
- Filo Lenarviricota
- Familia Botourmiaviridae
- Familia Narnaviridae
- Familia Mitoviridae
- Clase Leviviricetes
- Orden Norzivirales
- Familia Atkinsviridae
- Familia Duinviridae
- Familia Fiersviridae
- Familia Solspiviridae
- Orden Timlovirales
- Familia Blumeviridae
- Familia Steitzviridae
- Género Chimpevirus
- Género Hohglivirus
- Género Mahrahovirus
- Género Meihzavirus
- Género Nicedsevirus
- Género Sculuvirus
- Género Skrubnovirus
- Género Tetipavirus
- Género Winunavirus
- Orden Norzivirales
- Filo Duplornaviricota
- Familia Cystoviridae
- Familia Reoviridae
- Orden Ghabrivirales
- Familia Alternaviridae
- Familia Chrysoviridae
- Familia Megabirnaviridae
- Familia Quadriviridae
- Familia Totiviridae
- Filo Negarnaviricota
- Familia Tosoviridae
- Subfilo Haploviricotina
- Familia Qinviridae
- Familia Aspiviridae
- Familia Yueviridae
- Clase Monjiviricetes
- Orden Jingchuvirales
- Familia Chuviridae
- Familia Aliusviridae
- Familia Chuviridae
- Familia Crepuscuviridae
- Familia Myriaviridae
- Familia Nataviridae
- Orden Mononegavirales
- Familia Artoviridae
- Familia Bornaviridae
- Familia Filoviridae
- Familia Lispiviridae
- Familia Mymonaviridae
- Familia Nyamiviridae
- Familia Paramyxoviridae
- Familia Pneumoviridae
- Familia Rhabdoviridae
- Familia Sunviridae
- Familia Xinmoviridae
- Orden Jingchuvirales
- Subfilo Polyploviricotina
- Clase Ellioviricetes
- Orden Bunyavirales
- Familia Arenaviridae
- Familia Cruliviridae
- Familia Discoviridae
- Familia Fimoviridae
- Familia Hantaviridae
- Familia Leishbuviridae
- Familia Mypoviridae
- Familia Nairoviridae
- Familia Peribunyaviridae
- Familia Phasmaviridae
- Familia Phenuiviridae
- Familia Tospoviridae
- Familia Tulasviridae
- Familia Wupedeviridae
- Orden Bunyavirales
- Clase Ellioviricetes
- Clase Insthoviricetes
- Orden Articulavirales
- Familia Amnoonviridae
- Familia Orthomyxoviridae
- Orden Articulavirales
- Filo Kitrinoviricota
- Familia Flaviviridae
- Orden Nodamuvirales
- Familia Nodaviridae
- Familia Sinhaliviridae
- Orden Tolivirales
- Familia Carmotetraviridae
- Familia Tombusviridae
- Clase Alsuviricetes
- Orden Hepelivirales
- Familia Alphatetraviridae
- Familia Benyviridae
- Familia Hepeviridae
- Familia Matonaviridae
- Orden Martellivirales
- Familia Bromoviridae
- Familia Closteroviridae
- Familia Endornaviridae
- Familia Kitaviridae
- Familia Mayoraviridae
- Familia Togaviridae
- Familia Virgaviridae
- Orden Tymovirales
- Familia Alphaflexiviridae
- Familia Betaflexiviridae
- Familia Deltaflexiviridae
- Familia Gammaflexiviridae
- Familia Tymoviridae
- Orden Hepelivirales
- Filo Pisuviricota
- Clase Stelpaviricetes
- Familia Astroviridae
- Familia Potyviridae
- Clase Duplopiviricetes
- Orden Durnavirales
- Familia Amalgaviridae
- Familia Curvulaviridae
- Familia Hypoviridae
- Familia Fusariviridae
- Familia Partitiviridae
- Familia Picobirnaviridae
- Orden Durnavirales
- Clase Pisoniviricetes
- Orden Nidovirales
- Familia Coronaviridae
- Familia Mononiviridae
- Familia Tobaniviridae
- Suborden Adnidovirinae
- Familia Abyssoviridae
- Familia Arteriviridae
- Familia Cremegaviridae
- Familia Gresnaviridae
- Familia Olfoviridae
- Suborden Nanivirinae
- Familia Nanghosviridae
- Familia Nanhypoviridae
- Suborden Mesnidovirinae
- Familia Medioniviridae
- Familia Mesoniviridae
- Suborden Runidovirinae
- Familia Euroniviridae
- Familia Roniviridae
- Orden Picornavirales
- Familia Caliciviridae
- Familia Dicistroviridae
- Familia Iflaviridae
- Familia Marnaviridae
- Familia Noraviridae
- Familia Picornaviridae
- Familia Polycipiviridae
- Familia Secoviridae
- Familia Solinviviridae
- Orden Sobelivirales
- Familia Alvernaviridae
- Familia Barnaviridae
- Familia Solemoviridae
- Orden Yadokarivirales
- Familia Yadokariviridae
- Familia Hadakariviridae
- Familia Polymycoviridae
- Orden Nidovirales
- Familia Birnaviridae
- Familia Permutotetraviridae
- Clase Stelpaviricetes
- Filo Lenarviricota
- Familia Sarthroviridae
- Género Albetovirus
- Género Aumaivirus
- Género Papanivirus
- Género Virtovirus
- Reino Pararnavirae
- Dominio Adnaviria
- Familia Tristromaviridae
- Orden Ligamenvirales
- Familia Lipothrixviridae
- Familia Rudiviridae
- Familia Ungulaviridae
- Dominio Duplodnaviria
- Orden Herpesvirales
- Familia Alloherpesviridae
- Familia Herpesviridae
- Familia Malacoherpesviridae
- Clase Caudoviricetes
- Orden Crassvirales
- Familia Crevaviridae
- Familia Intestiviridae
- Familia Steigviridae
- Familia Suoliviridae
- Orden Kirjokansivirales
- Familia Flexireviridae
- Familia Haloferuviridae
- Familia Pyrstoviridae
- Familia Shortaselviridae
- Orden Juravirales
- Familia Yangangviridae
- Familia Yanlukaviridae
- Orden Methanobavirales
- Familia Anaeroviridae
- Familia Leisingerviridae
- Orden Nakonvirales
- Familia Ahpuchviridae
- Familia Ekchuahviridae
- Orden Thumleimavirales
- Familia Hafunaviridae
- Familia Halomagnusviridae
- Familia Queuoviridae
- Familia Soleiviridae
- Familia Aoguangviridae
- Familia Aliceevansviridae
- Familia Arenbergviridae
- Familia Ackermannviridae
- Familia Aggregaviridae
- Familia Assiduviridae
- Familia Autographiviridae
- Familia Casjensviridae
- Familia Chaseviridae
- Familia Demerecviridae
- Familia Drexlerviridae
- Familia Duneiviridae
- Familia Fervensviridae
- Familia Fredfastierviridae
- Familia Forsetiviridae
- Familia Herelleviridae
- Familia Helgolandviridae
- Familia Guelinviridae
- Familia Grimontviridae
- Familia Rountreeviridae
- Familia Salasmaviridae
- Familia Schitoviridae
- Familia Zobellviridae
- Familia Kleczkowskaviridae
- Familia Kyanoviridae
- Familia Madisaviridaeviridae
- Familia Molycoviridae
- Familia Pachyviridae
- Familia Peduoviridae
- Familia Pootjesviridae
- Familia Pungoviridae
- Familia Naomiviridae
- Familia Vilmaviridae
- Familia Zierdtviridae
- Familia Pervagoviridae
- Familia Orlajensenviridae
- Familia Mesyanzhinoviridae
- Familia Saparoviridae
- Familia Speroviridae
- Familia Straboviridae
- Familia Stanwilliamsviridae
- Familia Suolaviridae
- Familia Verdandiviridae
- Familia Vertoviridae
- Familia Winoviridae
- Familia Bartogtaviriformidae
- Familia Brachygtaviriformidae
- Familia Rhodogtaviriformidae
- Orden Crassvirales
- Orden Herpesvirales
- Dominio Monodnaviria
- Reino Loebvirae
- Familia Inoviridae
- Familia Plectroviridae
- Familia Paulinoviridae
- Reino Sangervirae
- Familia Microviridae
- Reino Trapavirae
- Familia Pleolipoviridae
- Reino Shotokuvirae
- Filo Cossaviricota
- Familia Bidnaviridae
- Familia Parvoviridae
- Clase Papovaviricetes
- Familia Papillomaviridae
- Familia Polyomaviridae
- Filo Creesdnaviricota
- Clase Repensiviricetes
- Familia Geminiviridae
- Familia Genomoviridae
- Clase Arfiviricetes
- Familia Bacilladnaviridae
- Familia Redondoviridae
- Familia Smacoviridae
- Familia Naryaviridae
- Familia Nenyaviridae
- Orden Cirlivirales
- Familia Circoviridae
- Familia Vilyaviridae
- Familia Anelloviridae
- Orden Mulpavirales
- Familia Nanoviridae
- Familia Amesuviridae
- Familia Metaxyviridae
- Familia Alphasatellitidae
- Clase Repensiviricetes
- Filo Cossaviricota
- Reino Loebvirae
- Dominio Varidnaviria
- Reino Helvetiavirae
- Orden Halopanivirales
- Familia Sphaerolipoviridae
- Familia Simuloviridae
- Familia Matshushitaviridae
- Familia Portogloboviridae
- Orden Halopanivirales
- Reino Bamfordvirae
- Filo Nucleocytoviricota
- Clase Megaviricetes
- Familia Mimiviridae
- Orden Algavirales
- Familia Phycodnaviridae
- Familia Yaraviridae
- Familia Mamonaviridae
- Orden Imitervirales
- Familia Mimiviridae
- Familia Allomimiviridae
- Familia Mesomimiviridae
- Familia Schizomimiviridae
- Orden Pimascovirales
- Familia Ascoviridae
- Familia Iridoviridae
- Familia Marseilleviridae
- Familia Pithoviridae
- Clase Pokkesviricetes
- Familia Asfarviridae
- Familia Poxviridae
- Clase Megaviricetes
- Filo Preplasmaviricota
- Familia Lavidaviridae
- Familia Adintoviridae
- Familia Finnlakeviridae
- Clase Tectiliviricetes
- Familia Adenoviridae
- Familia Tectiviridae
- Familia Turriviridae
- Orden Vinavirales
- Familia Corticoviridae
- Familia Autolykiviridae
- Clase Naldaviricetes
- Familia Nimaviridae
- Orden Lefavirales
- Familia Baculoviridae
- Familia Hytrosaviridae
- Familia Nudiviridae
- Familia Polydnaviriformidae
- Filo Nucleocytoviricota
- Reino Helvetiavirae
- Dominio Ribozyviria
- Familia Kolmioviridae
- Dominio Viroidia
- Familia Avsunviroidae
- Familia Pospiviroidae
- Familias de colocación incierta
- Dominio propuesto
- Familia Clavaviridae
- Virus fusiformes
- Familia Fuselloviridae
- Familia Halspiviridae
- Familia Thaspiviridae
- Familia Bicaudaviridae
- Familia Itzamnaviridae
- Familia Ampullaviridae
- Familia Globuloviridae
- Familia Guttaviridae
- Familia Plasmaviridae
- Familia Spiraviridae
- Familia Ovaliviridae
- Familia Tolecusatellitidae
- Dominio propuesto
- Género de colocación incierta
Sistema LHT de clasificación de virus
El Sistema LHT de Clasificación de Virus se basó en caracteres químicos y físicos como ácido nucleico (ADN o ARN), simetría (helicoidal o icosaédrica o compleja), presencia de envoltura, diámetro de la cápside , número de capsómeros. Esta clasificación fue aprobada por el Comité Provisional de Nomenclatura de Virus (PNVC) de la Asociación Internacional de Sociedades Microbiológicas (1962). Fue una clasificación taxonómica histórica y hoy en día ha sido remplazada por la del ICTV. La taxonomía fue la siguiente:
- Filo Vira (dividido en dos subfilos)
-
- Subfilo Deoxyvira (Virus ADN)
-
- Clase Deoxybinala
-
- Orden Urovirales
- Familia Phagoviridae
- Clase Deoxyhelica
-
- Orden Chitovirales
- Familia Poxviridae
- Clase Deoxycubica
-
- Orden Peplovirales
- Familia Herpesviridae
- Orden Haplovirales
- Familia Iridoviridae
- Familia Adenoviridae
- Familia Papiloviridae
- Familia Paroviridae
- Familia Microviridae
- Subfilo Ribovira (Virus ARN)
-
- Clase Ribocubica
-
- Orden Levivirales
- Familia Leviviridae
- Orden Togovirales
- Familia Arboviridae
- Orden Tymovirales
- Familia Cystoviridae
- Familia Napoviridae
- Familia Reoviridae
- Clase Ribohelica
-
- Orden Sagovirales
- Familia Stomataviridae
- Familia Paramyxoviridae
- Familia Myxoviridae
- Orden Rhabdovirales
-
- Suborden Flexiviridales
- Familia Mesoviridae
- Familia Peptoviridae
- Suborden Rigidovirales
- Familia Pachyviridae
- Familia Protoviridae
- Familia Polichoviridae
Véase también
Enlaces externos
-
 Wikispecies tiene un artículo sobre Clasificación de los virus.
Wikispecies tiene un artículo sobre Clasificación de los virus. - (en inglés) Sitio de la International Committee on Taxonomy of Viruses (ICTV)
- Lista de spp. versión 10 ICTV International Committee on Taxonomy of Viruses Master Species Lista 2009 (versión publicada el 24 de agosto de 2011) Archivado el 5 de marzo de 2016 en Wayback Machine.
- The Baltimore Method
- Virus Pathogen Database y Analysis Resource (ViPR)
- (en francés) Principios actuales de Clasificación de virus
- (en inglés) Índice oficial Los virus, después de ICTV, International Committee on Taxonomy of Viruses
- Base de datos de Taxonomía: Sitio NCBI Entrez Taxonomy