

La hipótesis de las dos corrientes es un modelo amplia mente aceptado e influyente del procesamiento neuronal de la visión y del oído. La hipótesis, dada su caracterización más popular en un artículo de Milner y Goodale en 1992, argumenta que los humanos poseen dos sistemas visuales distintos.
Recientemente, parece haber evidencia de dos sistemas auditivos distintos también.
A medida que la información visual sale del lóbulo occipital, y cuando el sonido sale de la red fonológica, sigue dos vías principales, o "corrientes". La corriente ventral (también conocido como "vía qué") está involucrado con la identificación y reconocimiento de objetos y visuales. La corriente dorsal o, ("vía dónde") está involucrada con el procesamiento de la ubicación espacial del objeto en relación con el espectador y con la repetición del habla.
Historia
Varios investigadores habían propuesto ideas similares anteriormente. Los propios autores acreditan la inspiración de trabajo en la visión ciega de Weiskrantz y la investigación de la visión de neurocientíficos anteriores. Schneider propuso por primera vez la existencia de dos sistemas visuales para localización e identificación en 1969. Ingle describió dos sistemas visuales independientes en ranas en 1973. Ettlinger revisó la evidencia neuropsicológica existente de una distinción en 1990. Además, Trevarthen había ofrecido una descripción de dos mecanismos de visión separados en monos en 1968.
En 1982, Ungerleider y Mishkin distinguieronlas corrientes dorsal y ventral, como procesamiento de características espaciales y visuales respectivamente, de sus estudios de lesiones de monos, proponiendo la distinction original del que y donde. Aunque este marco fue reemplazado por el de Milner y Goodale, sigue siendo influyente.
Una fuente de información enormemente influyente que ha informado al modelo ha sido el trabajo experimental que exploró las capacidades existentes del paciente agnóstico visual D.F. El primer informe, el más influyente, provino de Goodale y sus colegas en 1991 y todavía se está publicando un trabajo sobre ellas. Dos décadas después. Este ha sido el foco de algunas críticas del modelo debido a la dependencia excesiva percibida en los hallazgos de un solo caso.
Dos sistemas visuales
Goodale y Milner acumularon una serie de evidencias anatómicas, neuropsicológicas, electro fisiológicas y de comportamiento de su modelo. De acuerdo con sus datos, el flujo "perceptual" ventral calcula un mapa detallado del mundo a partir de la información visual, que luego se puede usar para operaciones cognitivas, y el flujo de "acción" dorsal transforma la información visual entrante al egocéntrico requerido (centrado en la cabeza) ) sistema de coordenadas para la planificación motriz especializada. El modelo también postula que la percepción visual codifica las propiedades espaciales de los objetos, como el tamaño y la ubicación, en relación con otros objetos en el campo visual; en otras palabras, utiliza métricas relativas y marcos de referencia basados en la escena. La planificación y coordinación de la acción visual, por otro lado, utiliza métricas absolutas determinadas a través de marcos de referencia egocéntricos, que calculan las propiedades reales de los objetos en relación con el observador. Por lo tanto, se ha demostrado que los movimientos de agarre dirigidos hacia objetos incrustados en escenas ambiguas de contraste de tamaño escapan a los efectos de estas ilusiones, ya que diferentes marcos de referencias y métricas están involucrados en la percepción de la ilusión contra la ejecución del acto de agarre.
Norman propuso un modelo de visión de proceso dual similar, y describió ocho diferencias principales entre los dos sistemas consistentes con otros modelos de dos sistemas.
| Factor | Sistema ventral | Sistema doral |
|---|---|---|
| Funcion | Reconocimiento/ identificación | Comportamiento guiado visualmente |
| Sensibilidad | Altas frecuencias espaciales-detalles | Frecuencias temporales altas-movimiento |
| Memoria | Representaciones almacenadas a largo plazo | Solo almacenamiento a muy corto plazo |
| Velocidad | Relativamente lento | Relativamente rápido |
| Conciencia | Típicamente alto | Típicamente bajo |
| Marco de referencia | Alocéntrico o centrado en el objeto | Egocéntrico o centrado en el espectador |
| Entrada visual | Principalmente foveal o parafoveal | A través de la retina |
| Vision monocular | Efectos generalmente razonablemente pequeños | A menudo grandes efectos, por ejemplo, paralaje de movimiento |
Corriente dorsal
Se propone que la corriente dorsal esté involucrada en la guía de acciones y reconocimiento de dónde están los objetos en el espacio. También conocida como la corriente parietal, la corriente "dónde" o la corriente "cómo", esta vía se extiende desde la corteza visual primaria (V1) en el lóbulo occipital hacia el lóbulo parietal. Está interconectada con la corriente ventral paralela (la corriente "qué") que corre hacia abajo desde V1 hasta el lóbulo temporal.
Características generales
La corriente dorsal está involucrada en la conciencia espacial y la orientación de las acciones (por ejemplo, alcance). En esto tiene dos características funcionales distintas: contiene un mapa detallado del campo visual y también es bueno para detectar y analizar movimientos.
La corriente dorsal comienza con funciones puramente visuales en el lóbulo occipital antes de pasar gradualmente a la conciencia espacial en su terminación en el lóbulo parietal.
La corteza parietal posterior es esencial para "la percepción e interpretación de las relaciones espaciales, la imagen corporal precisa y el aprendizaje de tareas que involucran la coordinación del cuerpo en el espacio".
Contiene lóbulos que funcionan individualmente. El surco parietal lateral (LIP) contiene neuronas que producen una activación mejorada cuando la atención se desplaza hacia el estímulo o las sacadas del animal hacia un estímulo visual, y el surco parietal (VIP) ventral donde se integra la información visual y sensorial.
Efectos del daño o lesiones.
El daño a la corteza parietal posterior causa una serie de trastornos espaciales que incluyen:
- Simultanagnosia: donde el paciente solo puede describir objetos individuales sin la capacidad de percibirlo como un componente de un conjunto de detalles u objetos en un contexto (como en un escenario, por ejemplo, el bosque para los árboles).
- Ataxia óptica : donde el paciente no puede usar información viso-espacial para guiar los movimientos del brazo.
- Desatención hemiespacial : cuando el paciente ignora la mitad contralesional del espacio (es decir, no se da cuenta de las cosas en su campo de visión izquierdo y se enfoca solo en los objetos en el campo de visión derecho; o no se da cuenta de las cosas en un campo de vista cuando los perciben en el otro). Por ejemplo, una persona con este trastorno puede dibujar un reloj y luego etiquetarlo de 12, 1, 2, ..., 6, pero luego detenerse y considerar que su dibujo está completo.
- Akinetopsia : incapacidad para percibir el movimiento.
- Apraxia : incapacidad para producir movimientos discrecionales o volitivos en ausencia de trastornos musculares.
Corriente ventral
El flujo ventral está asociado con el reconocimiento de objetos y la representación de formas. También se describe como la corriente "que", tiene fuertes conexiones con el lóbulo temporal medial (que almacena memoria a largo plazo ), el sistema límbico (que controla las emociones) y la corriente dorsal (que trata con la ubicación y el movimiento de los objetos).
La corriente ventral recibe su entrada principal de la capa parvo-celular (en oposición a la magnocelular ) del núcleo geniculado lateral del tálamo . Estas neuronas se proyectan a las subcapas V1 4Cβ, 4A, 3B y 2 / 3a sucesivamente. Desde allí, la vía ventral pasa por V2 y V4 hacia áreas del lóbulo temporal inferior : PIT (inferotemporal posterior), CIT (inferotemporal central) y AIT (inferotemporal anterior). Cada área visual contiene una representación completa del espacio visual. Es decir, contiene neuronas cuyos campos receptivos juntos representan todo el campo visual. La información visual entra en la corriente ventral a través de la corteza visual primaria (V1) y viaja a través del resto de las áreas en secuencia.
Moviéndose a lo largo del flujo de V1 a AIT, los campos receptivos aumentan su tamaño, latencia y la complejidad de su ajuste.
Todas las áreas en la corriente ventral están influenciadas por factores intraretinales además de la naturaleza del estímulo en su campo receptivo. Estos factores incluyen atención , memoria de trabajo y estímulo saliente. Por lo tanto, la corriente ventral no proporciona simplemente una descripción de los elementos en el mundo visual, sino que también desempeña un papel crucial al juzgar el significado de estos elementos.
El daño a la corriente ventral puede causar la incapacidad de reconocer caras o interpretar expresiones faciales
Dos sistemas auditivos
Corriente ventral

Junto con la vía ventral visual que es importante para el procesamiento visual, también hay una vía auditiva ventral que emerge de la corteza auditiva primaria. En esta vía, los fonemas se procesan posteriormente a sílabas y sonidos ambientales. La información luego se une a la corriente ventral visual en el giro temporal medio y el polo temporal. Aquí los objetos auditivos se convierten en conceptos audiovisuales interior.
Corriente dorsal
La función de la vía dorsal es mostrar representaciones sensoriales auditivas sobre representaciones motoras articulatorias. Hickok y Poeppel afirman que la vía dorsal es necesaria porque "aprender a hablar es esencialmente una tarea de aprendizaje motor. La entrada principal a esto es sensorial, el habla en particular. Por lo tanto, debe haber un mecanismo neuronal que codifique y mantenga instancias de sonidos del habla, y puede utilizar estos rastros sensoriales para guiar la sincronización de los gestos del habla para que los sonidos se reproducen con precisión ".
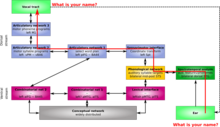
En contraste con el procesamiento auditivo de la corriente ventral, la información ingresa al oído y luego al giro temporal superior y, finalmente, al surco temporal superior. Desde allí, la información se desplaza al comienzo de la vía dorsal, que se encuentra en el límite de los lóbulos temporal y parietal cerca de la fisura de Sylvian. El primer paso de la vía dorsal comienza en la interfaz extrasensorial, ubicada en el parietal temporal izquierdo de Sylvian (Spt) (dentro de la fisura de Sylvian en el límite parietal-temporal). El spt es importante para percibir y reproducir sonidos. Esto es evidente porque su capacidad para adquirir nuevo vocabulario, ser interrumpida por lesiones y retroalimentación auditiva en la producción del habla, disminución articulatoria en la sordera tardía y el residuo no fonológico de la afasia de Wernicke; auto-control deficiente. También es importante para los mecanismos neuronales básicos de la memoria fonológica a corto plazo. Sin el Spt, la adquisición del lenguaje está deteriorada. La información luego pasa a la red articulatoria, que se divide en dos partes separadas. La red articulatoria 1, que procesa los programas de sílabas motoras, está ubicada en el giro temporal inferior posterior izquierdo y el área 44 de Brodmann (pIFG-BA44). La red articulatoria 2 es para programas de fonemas de motor y está ubicada en la izquierda M1-vBA6.
La afasia de conducción afecta la capacidad de un sujeto para reproducir el habla (generalmente por repetición), aunque no tiene influencia sobre la capacidad del sujeto para comprender el lenguaje hablado. Esto demuestra que la afasia de conducción no debe reflejar un deterioro de la vía ventral sino la dorsal. Buchsbaum et al encontraron que la afasia de conducción puede ser el resultado de un daño, en particular de lesiones, en el Spt (parietal temporal de Sylvian). Esto se demuestra por la participación del Spt en la adquisición de nuevo vocabulario, ya que aunque los experimentos han demostrado que la mayoría de los afasias de conducción pueden repetir palabras simples y de alta frecuencia, su capacidad para repetir palabras complejas de baja frecuencia se ve afectada. El Spt es responsable de conectar el motor y los sistemas auditivos al hacer que el código auditivo sea accesible para la corteza del motor. Parece que la corteza motora recrea palabras simples de alta frecuencia (como cup) para acceder a ellas de manera más rápida y eficiente, mientras que las palabras complejas de baja frecuencia (como Sylvian parietal temporal) requieren una regulación en línea más activa por parte del SPT. Esto explica por qué las afasias de conducción tienen una dificultad particular con las palabras de baja frecuencia que requieren un proceso más práctico para la producción del habla. "La afasia de conducción se ha caracterizado como un déficit en la capacidad de codificar la información fonológica para la producción", principalmente debido a una interrupción en la interfaz motor-auditiva. La afasia de conducción se ha relacionado más específicamente con el daño del folículo arqueado, que es vital tanto para el habla como para la comprensión del lenguaje, ya que el folículo arqueado constituye la conexión entre las áreas de Broca y Wernicke.
Críticas
La innovación de Goodale & Milner fue cambiar la perspectiva de un énfasis en las distinciones de entrada, como la ubicación del objeto frente a las propiedades, a un énfasis en la relevancia funcional de la visión para el comportamiento, la percepción o la acción. Sin embargo, las perspectivas contemporáneas, informadas por el trabajo empírico en las últimas dos décadas, ofrecen una explicación más compleja que una simple separación de funciones en dos corrientes. El trabajo experimental reciente, por ejemplo, ha cuestionado estos hallazgos y ha sugerido que la aparente disociación entre los efectos de las ilusiones sobre la percepción y la acción se debe a las diferencias en la atención, las demandas de tareas y otros factores de confusión. Sin embargo, hay otros hallazgos empíricos que no se pueden descartar tan fácilmente, lo que brinda un fuerte apoyo a la idea de que las acciones expertas no se ven afectadas por las ilusiones pictóricas.
Además, las investigaciones neuropsicológicas recientes han cuestionado la validez de la disociación de las dos corrientes que ha proporcionado la piedra angular de la evidencia para el modelo. La disociación entre la agnosia visual y la ataxia óptica ha sido cuestionada por varios investigadores por no ser tan fuerte como se describió originalmente; Hesse y sus colegas demostraron deficiencias en la corriente dorsal en el DF del paciente; Himmelbach y sus colegas revaluaron las habilidades del DF y aplicaron un análisis estadístico más riguroso que demuestra que la disociación no fue tan fuerte como se pensaba al principio.
Una revisión de 2010 de la evidencia acumulada para el modelo concluyó que, si bien el espíritu del modelo ha sido reivindicado, la independencia de las dos corrientes se ha exagerado. Goodale y Milner han propuesto la analogía de la tele asistencia, uno de los esquemas más eficientes diseñados para el control remoto de robots que trabajan en entornos hostiles. En esta cuenta, el flujo dorsal se ve como una función semiautomatica que opera bajo la guía de las funciones ejecutivas que a su vez se informan mediante el procesamiento del flujo ventral.
Así, la perspectiva emergente dentro de la neuropsicología y la neurofisiología es que, si bien un marco de dos sistemas fue un avance necesario para estimular el estudio de las funciones altamente complejas y diferenciadas de las dos vías neuronales; es más probable que la realidad implique una interacción considerable entre la visión para la acción y la visión para la percepción. Rob McIntosh y Thomas Schenk resumen esta posición de la siguiente manera:
Debemos ver el modelo no como una hipótesis formal, sino como un conjunto de heurísticas para guiar el experimento y la teoría. Los diferentes requisitos informativos del reconocimiento visual y la guía de acción aún ofrecen una explicación convincente para las amplias especializaciones relativas de las corrientes dorsal y ventral. Sin embargo, para avanzar en el campo, es posible que tengamos que abandonar la idea de que estas corrientes funcionan en gran medida independientemente de las demás, y abordar los detalles dinámicos de cómo las muchas áreas del cerebro visual se organizan de una tarea a otra en redes funcionales novedosas.
Enlaces externos
- Esta obra contiene una traducción total derivada de «Two-streams hypothesis» de Wikipedia en inglés, concretamente de esta versión, publicada por sus editores bajo la Licencia de documentación libre de GNU y la Licencia Creative Commons Atribución-CompartirIgual 3.0 Unported.
| Control de autoridades |
|
|---|
-
 Datos: Q946325
Datos: Q946325
-
 Multimedia: Ventral and dorsal stream / Q946325
Multimedia: Ventral and dorsal stream / Q946325