
En biología se denomina especiación al proceso mediante el cual una población de una determinada especie da lugar a otra u otras especies. El proceso de especiación, a lo largo de 3.800 millones de años, ha dado origen a una enorme diversidad de organismos, millones de especies de todos los reinos, que han poblado y pueblan la Tierra casi desde el momento en que se formaron los primeros mares.
A pesar de que el aislamiento geográfico juega un papel importante en la mayoría de los casos de especiación no es el único factor.
El término se aplica a un proceso de división de clados (cladogénesis) más que el de evolución de una especie a otra (anagénesis). La especiación también puede ocurrir artificialmente en la cría de animales, plantas e incluso en experimentos de laboratorio.
Charles Darwin fue el primero en escribir sobre el papel de la selección natural en la especiación. Existen investigaciones sobre los efectos que tiene la selección sexual sobre la especiación pero todavía no se han podido confirmar definitivamente si esta es una de las causas de la especiación o no.
Principales mecanismos de especiación
Cladogénesis
La cladogénesis o bifurcación es el mecanismo de especiación más importante. Se produce por aislamiento reproductivo de diferentes poblaciones de una especie debido a las barreras a la hibridación que pueden ser precigóticas o postcigóticas.
- Las barreras precigóticas son mecanismos de aislamiento que tienen lugar antes o durante la fecundación, a la que limitan, actúan antes del intercambio gamético. Puede ser por aislamiento ecológico, etológico o mecánico.
- Las barreras postcigóticas son todas las que atañen a la viabilidad de los individuos producidos, a través de abortos espontáneos, esterilidad del híbrido, muerte prematura, híbridos débiles y enfermizos, etc.
Hibridación
Hibridación es el cruce reproductivo entre dos especies distintas que pueden producir individuos viables, que pueden ser o no fértiles. Aunque la hibridación en animales puede ser un proceso natural, lo más frecuente es que esté asociado a alteraciones provocadas por la introducción de nuevas especies por el hombre desde que descubrió la ganadería. Es sumamente difícil que esto dé lugar a nuevas especies en los animales.
Entre las plantas la hibridación es un fenómeno extremadamente común. La introgresión es la penetración de genes de una especie en otra por medio de la hibridación. Los híbridos tienden a derivar, en las sucesivas generaciones, hacia los caracteres de uno de los parentales, pero en el proceso puede quedar fijada la transferencia de algunos genes desde la otra especie. No obstante, al igual que en los animales la formación de nuevas especies por hibridación es sumamente rara en las plantas.
Tipos de especiación

Especiación geográfica o alopátrica
La especiación geográfica o alopátrica es el modelo que cuenta con un mayor número de ejemplos ampliamente documentados. Básicamente es la especiación gradual que se produce cuando una especie ocupa una gran área geográfica que no permite que los individuos que estén muy alejados puedan cruzarse entre sí, debido a barreras geográficas como mares, montañas o desiertos. Se trata, entonces, de la separación geográfica de un acervo genético continuo, de tal forma que se establecen dos o más poblaciones geográficas aisladas. La separación entre las poblaciones puede ser debida a migración, a extinción de las poblaciones situadas en posiciones geográficas intermedias, o mediada por sucesos geológicos. La barrera puede ser geográfica o ecológica, como por ejemplo cumbres que separan valles en las cordilleras o zonas desérticas que separan zonas húmedas. La separación espacial de dos poblaciones de una especie durante un largo periodo de tiempo da lugar a la aparición de novedades evolutivas en una o en las dos poblaciones debido a que el medio ambiente es distinto en las diferentes zonas geográficas; se detiene el flujo genético entre poblaciones. Este tipo de especiación puede a su vez ser dividido en dos clases:
En el modelo I (especiación geográfica o vicariante) la especiación se produce por la separación de una especie ancestral en dos poblaciones relativamente grandes que permanecen aisladas al menos hasta la aparición de independencia evolutiva. La especiación es producto de procesos microevolutivos que producen divergencia gracias a la adaptación local y la diferenciación geográfica. La diferenciación puede ser debida tanto a factores estocásticos (deriva genética) como a procesos selectivos (adaptación). Sin embargo, puesto que se supone que el tamaño de población no es pequeño, los factores estocásticos no serán los más importantes en este modelo de especiación.
Quizás el ejemplo mejor documentado de este tipo de especiación es el producido por la formación del Istmo de Panamá hace aproximadamente tres millones de años. La aparición de esta barrera geográfica separó las poblaciones de organismos acuáticos en lo que actualmente son aguas de los océanos Pacífico y Atlántico. Dentro del género de langostas Alpheus se han hallado siete parejas de especies muy estrechamente relacionadas, con un representante a cada lado del Istmo de Panamá. Estas son consideradas especies gemelas que difieren ligeramente en morfología y que están aisladas genéticamente tanto por características etológicas como por barreras postcigóticas. Las evidencias moleculares (tanto isoenzimas como DNA) indican que la divergencia se ha producido tras la aparición del Istmo de Panamá. En general, los animales grandes y de gran movilidad (e.g., vertebrados carnívoros, ciertas aves, peces) son candidatos para especiar por este tipo de especiación geográfica. No obstante, se ha citado este mecanismo de especiación en hongos, moluscos braquiópodos fósiles.
El modelo II recoge el tipo de especiación alopátrica aplicable a especies asexuales, donde no hay evidencias de flujo genético entre poblaciones y donde la identidad como especie está determinada por motivos históricos (e.g., estasis evolutiva). En estos casos, la deriva genética puede ser el factor evolutivo más importante y la especiación puede ser rápida. Pero si las poblaciones originales están sometidas a variación geográfica, la selección debida a presiones adaptativas locales también puede ser un factor importante en la especiación.
Especiación peripátrica
En la especiación mediante poblaciones periféricas o especiación peripátrica, la nueva especie surge en hábitats marginales, habitualmente en los límites de distribución de una población central de menor tamaño. El flujo interdémico entre estas poblaciones puede reducirse y finalmente ser inexistente, gracias a lo cual estas poblaciones periféricas pueden convertirse en especies diferentes. Estaríamos ante casos de aislamiento geográfico, y posterior especiación, producidos por fenómenos de dispersión y colonización. Igual que en el caso anterior, la adaptación a esos ambientes puede ser el desencadenante de la divergencia, pero al tratarse de poblaciones pequeñas, los factores estocásticos pueden tener mayor importancia. Como ejemplos de este tipo de especiación podemos considerar la radiación evolutiva de las especies de Drosophila en Hawái. El amplio número de especies de drosophilidos en este archipiélago (más de 500 especies), parece haber surgido mediante especiación por migración y diversificación. Muchas de estas especies son endémicas de una isla, lo que sugiere que son productos de la fundación de una colonia por muy pocos individuos, provenientes de una isla próxima, y de la posterior divergencia evolutiva. Varias de las predicciones de este modelo (las especies emparentadas deben estar en islas próximas, y las especies más modernas en islas formadas más recientemente) han sido confirmadas mediante técnicas moleculares. Los organismos que presentan una menor capacidad de dispersión y que se estructuran en pequeñas poblaciones son, en general, los mejores candidatos para este tipo de especiación.
Especiación simpátrica o simpátrida
La especiación simpátrica implica la divergencia de algunos demos (poblaciones locales) hasta conseguir independencia evolutiva dentro de un mismo espacio geográfico. Habitualmente conlleva que las nuevas poblaciones utilicen nichos ecológicos diferentes, dentro del rango de distribución de la especie ancestral, desarrollando mecanismos de aislamiento reproductivo. La divergencia en simpatría puede estar impulsada por la especialización ecológica de algunos demes, aunque también existe la posibilidad de que la especiación se produzca por hibridación entre especies muy próximas. Otro tipo de especiación simpátrica implica la formación de una especie asexual a partir de una especie precursora sexual. El escenario teórico más habitual asume selección disruptiva implicando al menos dos loci: uno (A) para la adaptación a un recurso y un segundo (C) que rige el comportamiento de un organismo, como por ejemplo un insecto que se alimenta de una planta. La especie ancestral pongamos por caso que tuviera genotipo A1A1, el cual le confiere adaptación a consumir eficazmente cierta planta “uno”. Una mutación A2 confiere la capacidad de consumir eficazmente la planta “dos”, al menos cuando aparece en homocigosis, pero al mismo tiempo reduce la adaptación a la planta 1. De igual forma, los homocigotos A1 presentan una menor adaptación frente a la planta 2. Los heterocigotos A1A2 presentan una menor eficacia biológica en cualquier planta. La selección favorecerá el incremento de A2, aunque debido al apareamiento aleatorio se producirán heterocigotos de menor eficacia biológica. Si el locus C con dos alelos (C1 y C2) controla el apareamiento, y estos alelos producen apareamiento clasificado positivo (esto es, los apareamientos se producen entre individuos de igual genotipo para este locus), se dan las condiciones para que puedan aparecer combinaciones A1A1 C1C1 y A2A2 C2C2 que preferirán individuos similares para aparearse y tendrán mayor eficacia biológica en sus respectivos recursos preferidos (plantas 1 y 2). El modelo plantea que si las diferencias en eficacia biológica fuesen amplias y estos genes estuvieran en desequilibrio de ligamiento (ligados), se podría producir el aislamiento etológico necesario para que se produzca la especiación. Por el contrario, la recombinación rompe estas combinaciones y es la fuerza que impide que se desarrolle el aislamiento reproductivo.
Una variante de este modelo implica que el locus C controle la “preferencia de hospedador” y que el apareamiento se produzca en la planta preferida. En esta situación los individuos C1 preferirán la planta 1, y los C2 la planta 2. Los mutantes C2 serían seleccionados puesto que hacen uso del recurso menos utilizado (reduciendo así la competencia intraespecífica). Si C2 se asocia con A2 se dan las condiciones para un verdadero aislamiento ecológico, ya que los apareamientos entre homocigotos diferentes serán poco frecuentes y el número de progenie heterocigota se reducirá. Este modelo es más robusto en el sentido de que no es necesario un gran desequilibrio de ligamiento entre ambos loci para que se produzcan dos variedades aisladas ecológicamente. La mosca de la fruta Rhagoletis pomonella representa un claro ejemplo del anterior modelo teórico. Estas moscas se aparean sobre la superficie de las frutas de algunas especies vegetales (el hospedador) y, tras la ovoposición, las larvas se desarrollan dentro de la fruta. Varias especies dentro del género Rhagoletis han cambiado de especie hospedadora recientemente. Rhagoletis pomonella pasó de utilizar su hospedador habitual, un espino del género Crataegus, a utilizar una especie introducida, el manzano, en 1864 en el valle del río Hudson (Estados Unidos). En Wisconsin, han aparecido razas de la misma especie que han pasado de utilizar el manzano a preferir el cerezo. En otras especies del mismo género, se han producido cambios de hospedador similares.
La especiación simpátrica parece haber sido frecuente entre las especies parásitas tanto de animales como de plantas y entre los insectos fitófagos, donde la especialización puede haber sido el factor desencadenante de la especiación. También diversas especies de peces y organismos acuáticos que habitan lagos parecen ser candidatos típicos a fenómenos de especiación simpátrica. La gran diversidad en especies de peces cíclidos que habitan lagos africanos situados en antiguos cráteres parece haber surgido por especiación simpátrica, quizá propiciada por apareamiento clasificado, según parecen indicar las filogenias de DNA. Estas filogenias indican que las especies que habitan un mismo lago son monofiléticas, implicando sucesos de especiación simpátrica en un mismo entorno, donde no existen grandes diferencias en microhábitats que pudieran provocar situaciones de micro-alopatría.
Si dos especies recientemente originadas, y con un aislamiento genético aún no completado totalmente, entran en contacto (contacto secundario) pueden hibridar. Estos híbridos pueden ser de baja eficacia biológica o, por el contrario, mostrar rasgos característicos que sean ventajosos frente a las especies parentales. Así se pueden crear zonas híbridas, donde según ciertos criterios de especie, se identifican a estos híbridos como nuevas especies. Incluso algunos de los híbridos pueden desarrollar independencia evolutiva y ser considerados como especies bajo todos los criterios. En general, la especiación por hibridación es un tipo de especiación ampliamente encontrada entre los vegetales.
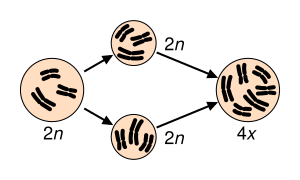
Los híbridos entre dos especies necesitan tener coadaptados los dos genomas de los que proceden, lo que implica que este tipo de especiación se reserve a especies emparentadas. Quizás las diferencias en sistema de reproducción sea el problema principal que deben afrontar los híbridos en el camino hacia la estabilización de sus genomas, y por lo tanto hacia la especiación por hibridación. La reproducción vegetativa, la agamospermia (producción de semillas sin necesidad de fecundación), la anfiploidía y la poliploidía han sido las vías más utilizadas por diversas especies vegetales originadas por hibridación para estabilizar sus genomas y obtener independencia evolutiva de sus especies ancestrales. La reproducción vegetativa y la agamospermia han permitido a ciertas plantas híbridas reproducirse sin hacer uso de la meiosis, que en los híbridos suele ser anormal al no aparear correctamente los cromosomas de las especies progenitoras. Ambos procesos permiten que se formen especies híbridas, que habitualmente presentan ámbitos de distribución muy limitados (microespecies). El tipo de especiación híbrida más frecuente entre plantas parece haber sido la anfiploidía o alopoliploidía. Al menos la mitad de todos los poliploides naturales son de origen alopoliploide. Este proceso implica la duplicación de todos los cromosomas de un híbrido para dar lugar a un individuo con un mayor grado de ploidía. Si una especie de genoma AA y 2n=10 hibrida con otra especie de genoma BB y 2n=12, el híbrido poseerá una composición genética AB y 11 cromosomas. Tras el proceso de endorreduplicación de los cromosomas pasará a tener 22 cromosomas y ser AABB, con lo cual cada cromosoma tendrá su homólogo, se evitan problemas meióticos derivados de un mal apareamiento cromosómico -que habitualmente conducen a la esterilidad- y se restablece la reproducción sexual. Como ejemplo, se puede citar a la variedad “Golden yellow“ del azafrán de flor Crocus flavus. Esta variedad es realmente una especie (2n=14) originada por anfiploidía al hibridar C. flavus (2n=8) y C. angustifolius (2n=6).
El surgimiento de plantas asexuales o autógamas a partir de poblaciones sexuales y alógamas conduce inmediatamente a la independencia evolutiva y puede considerarse un tipo de especiación simpátrica instantánea. La reproducción asexual puede producirse de diversas formas, básicamente mediante la generación de nuevos organismos a partir de células somáticas (reproducción vegetativa), mediante el desarrollo de semillas a partir de óvulos no fecundados (apomixis) o mediante el desarrollo a partir del óvulo, con la necesidad del gameto masculino, pero sin que este aporte material genético (pseudogamia). La mayoría de las especies que utilizan estos sistemas, al menos en plantas, proceden de eventos de hibridación. Una vez que se ha establecido una especie de este tipo, los individuos pueden divergir vía mutaciones somáticas, formando grupos de microespecies. Las especies asexuales aparecen tanto en animales como en plantas, comprendiendo, algunos lagartos, salamandras, peces, planarias e insectos, pero sobre todo angiospermas y algunos helechos. La autogamia produce exactamente el mismo efecto de aislamiento con respecto a la población original por lo que se le considera otro factor en la especiación saltacional. Este tipo de reproducción permite que produzcan descendientes aunque no existan otros individuos de la misma especie en el mismo sitio (por ejemplo, en el caso de especies colonizadoras), o que los polinizadores sean escasos (en el caso de las especies zoófilas). Un ejemplo de este tipo de especiación es la del teporingo (Romerolagus diazi).
Especiación instantánea o cuántica
La especiación cuántica, también llamada saltacional, es el proceso por el cual una población pequeña de una especie diverge rápidamente en más de una especie que se halla reproductivamente aislada de la población original.
Especiación parapátrica
Según este modelo, la especiación se produce sin una separación geográfica completa de las poblaciones. De hecho, la especie hija puede compartir parte del rango de distribución con la especie madre e hibridar en las zonas de contacto. Los demás suelen divergir debido tanto a factores estocásticos como a selección local. La migración inter-démica y la hibridación pueden contrarrestar la divergencia, pero en determinadas circunstancias, la selección diversificadora (a favor de variaciones locales) puede ser el factor más importante, implicando la formación de clinas en la frecuencia de algunos loci. El apareamiento clasificado y una reducción en eficacia biológica de los híbridos conduciría al desarrollo final de la especiación.
En general, este tipo de especiación es difícilmente distinguible de la especiación alopátrica seguida de un contacto secundario posterior, lo cual ha conducido a amplios debates acerca de la importancia (e incluso realidad) de este tipo de especiación. La planta Anthoxanthum odoratum podría representar un candidato a este tipo de especiación. Varias poblaciones de esta especie han desarrollado tolerancia a metales pesados en zonas contaminadas con estas sustancias. Los metales pesados representan un factor selectivo muy importante en estas áreas, y las plantas no tolerantes no prosperan adecuadamente. Las poblaciones tolerantes han empezado a divergir frente a la especie “parental” no solo en su capacidad de soportar altas concentraciones de estos metales, sino también en características fenológicas y en capacidad de autofecundación. Estas diferencias con las poblaciones limítrofes no tolerantes implican la aparición de mecanismos de aislamiento reproductivo de forma parapátrica.
Las plantas, los moluscos terrestres, pequeños mamíferos, insectos no voladores y algunos anfibios y reptiles, son los organismos candidatos a este tipo de especiación. En general, se trata de organismos con hábitos sedentarios y con poca capacidad de dispersión.
Un caso intermedio entre la especiación alopátrica y la parapátrica es la denominada alo-parapátrica. Básicamente la situación de especiación es alopátrica (modelo I), pero en este caso, las poblaciones entran en contacto antes de que se haya producido una divergencia completa (y la aparición de barreras de aislamiento eficientes). La especiación puede producirse tras un periodo de contacto, donde el reforzamiento podría ser un factor determinante.
La especiación estasipátrica se ha considerado sinónima de la anterior y también como un tipo especial de la especiación alopátrica, implicando la aparición de independencia evolutiva debida a mutaciones cromosómicas. Este tipo de especiación puede aparecer cuando se produce una mutación o reordenación cromosómica (e.g., inversión pericéntrica, fusión o translocación) que posibilita a los individuos portadores colonizar con mayor éxito un hábitat o área contigua al de la especie “madre”. Estas reordenaciones hacen que los individuos heterocigotos sean inviables o tengan su eficacia biológica muy reducida al no estar adaptados a ninguno de los hábitat, ni al original ni al nuevo. Estas mutaciones pueden ser fijadas por deriva, consanguinidad o acumulación meiótica en pequeñas poblaciones con poca migración, dando lugar a una nueva especie que se sitúa dentro del mismo rango de distribución que la especie original, en situaciones de parapatría. Las nuevas especies serán genéticamente muy parecidas a la especie ancestral, diferenciándose en alguna característica cariológica, para la cual estarán en condición homozigótica.
Los saltamontes australianos del género Vandiemenella, compuesto por más de 240 especies, pueden ilustrar este tipo de especiación. Todas estas especies presentan distribuciones parapátricas, siendo raras las zonas de simpatría. La distribución de especies parece ser estable desde el pleistoceno, contribuyendo a ello la poca movilidad de estos saltamontes no alados. El amplio número de especies puede ser debido a que éstas presentan cariotipos exclusivos con un amplio espectro de variaciones cromosómicas estructurales interespecíficas.
Véase también
Enlaces externos
- [1]
- 80 años....."todo alopátrico..salvo estos ejemplos que son la punta del iceberg...."
- [2] (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).* especciación híbrida
- Homoploid Reticulate Evolution in Helianthus (Asteraceae): Evidence from Ribosomal Genes
Loren H. Rieseberg American Journal of Botany, Vol. 78, No. 9 (Sep., 1991), pp. 1218-1237
- El contenido de este artículo incorpora material de una entrada de la Enciclopedia Libre Universal, publicada en español bajo la licencia Creative Commons Compartir-Igual 3.0.
| Control de autoridades |
|
|---|
-
 Datos: Q39350
Datos: Q39350
-
 Multimedia: Speciation / Q39350
Multimedia: Speciation / Q39350