
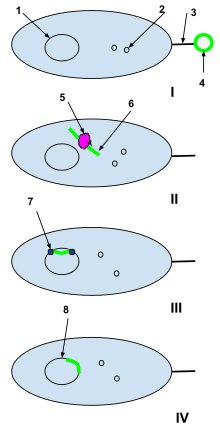
1-ADN de células bacterianas
2-Plásmidos de células bacterianas
3-Pili de sexos
4-Plásmido de ADN extraño de una célula muerta
5-Enzima de restricción de células bacterianas
6-Plásmido extraño desenrollado
7-ADN ligasa
I: Un plásmido de ADN extraño de una célula muerta es interceptado por los pili sexuales de una célula bacteriana naturalmente competente.
II: El plásmido extraño se transduce a través de los pili sexuales hacia la célula bacteriana, donde es procesado por enzimas de restricción de células bacterianas. Las enzimas de restricción rompen el plásmido extraño en una cadena de nucleótidos que se pueden agregar al ADN bacteriano.
III: la ADN ligasa integra los nucleótidos extraños en el ADN de la célula bacteriana.
IV: La recombinación está completa y el ADN extraño se ha integrado en el ADN de la célula bacteriana original y seguirá siendo parte de él cuando la célula bacteriana se replique a continuación.
En microbiología, genética, biología celular y biología molecular, la competencia es la capacidad de una célula para alterar su genética al absorber ADN extracelular ("desnudo") de su entorno en el proceso llamado transformación. La competencia se puede diferenciar entre competencia natural, una capacidad genéticamente especificada de las bacterias que se cree que ocurre en condiciones naturales así como en el laboratorio, y competencia inducida o artificial, que surge cuando las células en cultivos de laboratorio se tratan para hacerlas transitoriamente permeables a ADN. La competencia permite una rápida adaptación y reparación del ADN de la célula. Este artículo trata principalmente de la competencia natural en bacterias, aunque también se proporciona información sobre la competencia artificial.
Historia
La competencia natural fue descubierta por Frederick Griffith en 1928, cuando demostró que una preparación de células muertas de una bacteria patógena contenía algo que podía transformar células no patógenas relacionadas en el tipo patógeno. En 1944 Oswald Avery, Colin MacLeod y Maclyn McCarty demostraron que este "factor transformador" era ADN puro. Esta fue la primera evidencia convincente de que el ADN transporta la información genética de la célula.
Desde entonces, se ha estudiado la competencia natural en varias bacterias diferentes, en particular Bacillus subtilis, Streptococcus pneumoniae ("neumococo" de Griffith), Neisseria gonorrhoeae, Haemophilus influenzae y miembros del género Acinetobacter. Las áreas de investigación activa incluyen los mecanismos de transporte de ADN, la regulación de la competencia en diferentes bacterias y la función evolutiva de la competencia.
Mecanismos de captación de ADN
En el laboratorio, el investigador proporciona el ADN, a menudo como un fragmento o plásmido modificado genéticamente. Durante la captación, el ADN se transporta a través de la membrana celular y la pared celular, si está presente. Una vez que el ADN está dentro de la célula, puede degradarse a nucleótidos, que se reutilizan para la replicación del ADN y otras funciones metabólicas. Alternativamente, puede recombinarse en el genoma de la célula mediante sus enzimas de reparación del ADN. Si esta recombinación cambia el genotipo de la célula, se dice que la célula se ha transformado. La competencia y la transformación artificiales se utilizan como herramientas de investigación en muchos organismos (consulte Transformación (genética)).
En casi todas las bacterias naturalmente competentes, los componentes de los filamentos extracelulares llamados pili tipo IV (un tipo de fimbria) se unen al ADN extracelular de doble hebra. Luego, el ADN se transloca a través de la membrana (o membranas para bacterias gram negativas) a través de complejos de proteínas de componentes múltiples impulsados por la degradación de una hebra del ADN. El ADN monocatenario de la célula está unido por una proteína bien conservada, DprA, que carga el ADN en RecA, que media la recombinación homóloga a través de la ruta clásica de reparación del ADN.
Regulación de competencia
En cultivos de laboratorio, la competencia natural suele estar estrictamente regulada y, a menudo, se desencadena por escasez nutricional o condiciones adversas. Sin embargo, las señales inductoras específicas y la maquinaria reguladora son mucho más variables que la maquinaria de captación, y se sabe poco sobre la regulación de la competencia en los entornos naturales de estas bacterias. Se han descubierto factores de transcripción que regulan la competencia; un ejemplo es sxy (también conocido como tfoX) que se ha encontrado regulado a su vez por un elemento de ARN no codificante 5 '. En las bacterias capaces de formar esporas, las condiciones que inducen la esporulación a menudo se superponen con las que inducen la competencia. Por tanto, los cultivos o colonias que contienen células esporulantes a menudo también contienen células competentes. Investigaciones recientes han identificado un módulo central excitable de genes que puede explicar la entrada y salida de la competencia cuando se tiene en cuenta el ruido celular.
Se cree que la mayoría de las bacterias competentes absorben todas las moléculas de ADN con eficiencias aproximadamente iguales, pero las bacterias de las familias Neisseriaceae y Pasteurellaceae toman preferentemente fragmentos de ADN que contienen secuencias cortas de ADN, lo que se denomina secuencia de absorción de ADN (DUS) en Neisseriaceae y secuencia de señal de absorción (USS) en Pasteurellaceae, que son muy frecuentes en sus propios genomas. Los genomas de Neisserial contienen miles de copias de la secuencia GCCGTCTGAA, y los genomas de Pasteurellacean contienen AAGTGCGGT o ACAAGCGGT.
Funciones evolutivas y consecuencias de la competencia
La mayoría de las hipótesis propuestas para la función evolutiva primaria de la competencia natural como parte de la transformación bacteriana natural se dividen en tres categorías:
- la ventaja selectiva de la diversidad genética
- absorción de ADN como fuente de nucleótidos (ADN como "alimento")
- la ventaja selectiva de una nueva hebra de ADN para promover la reparación recombinacional homóloga del ADN dañado (reparación del ADN)
También se ha hecho una sugerencia secundaria, señalando la ventaja ocasional de la transferencia horizontal de genes.
Hipótesis de diversidad genética
Existen argumentos para apoyar la diversidad genética como la principal función evolutiva del sexo (incluida la transformación bacteriana). Sin embargo, las dificultades teóricas asociadas con la evolución del sexo sugieren que el sexo para la diversidad genética es problemático. Específicamente con respecto a la transformación bacteriana, la competencia requiere el alto costo de un interruptor de síntesis de proteínas global, con, por ejemplo, más de 16 genes que se encienden solo durante la competencia de Streptococcus pneumoniae. Sin embargo, dado que las bacterias tienden a crecer en clones, el ADN disponible para la transformación generalmente tendría el mismo genotipo que el de las células receptoras. Por tanto, siempre existe un coste elevado en la expresión de proteínas sin, en general, un aumento de la diversidad. Se han considerado otras diferencias entre competencia y sexo en modelos de evolución de genes que causan competencia; Estos modelos encontraron que los beneficios recombinacionales postulados de la competencia eran incluso más esquivos que los del sexo.
Hipótesis del ADN como alimento
La segunda hipótesis, el ADN como alimento, se basa en el hecho de que las células que absorben el ADN inevitablemente adquieren los nucleótidos que componen el ADN y, dado que los nucleótidos son necesarios para la síntesis de ADN y ARN y son costosos de sintetizar, estos pueden hacer una contribución significativa al presupuesto de energía de la célula. Algunas bacterias naturalmente competentes también secretan nucleasas en su entorno, y todas las bacterias pueden absorber los nucleótidos libres que estas nucleasas generan a partir del ADN ambiental. La energía de la absorción de ADN no se comprende en ningún sistema, por lo que es difícil comparar la eficiencia de la secreción de nucleasas con la de la absorción de ADN y la degradación interna. En principio, el costo de producción de nucleasas y la incertidumbre de la recuperación de nucleótidos deben equilibrarse con la energía necesaria para sintetizar la maquinaria de absorción y atraer el ADN. Otros factores importantes son la probabilidad de que las nucleasas y las células competentes encuentren moléculas de ADN, las ineficiencias relativas de la captación de nucleótidos del medio ambiente y del periplasma (donde una hebra es degradada por las células competentes) y la ventaja de producir moléculas listas para usar monofosfatos de nucleótidos de la otra hebra en el citoplasma. Otro factor de complicación es el sesgo propio de los sistemas de captación de ADN de las especies de la familia Pasteurellaceae y el género Neisseria, que podría reflejar la selección para la recombinación o la captación mecánicamente eficiente.
Hipótesis de reparación del daño del ADN
En las bacterias, el problema del daño del ADN es más pronunciado durante los períodos de estrés, particularmente el estrés oxidativo, que se produce durante las condiciones de hacinamiento o inanición. En tales condiciones, a menudo hay un solo cromosoma presente. El hallazgo de que algunas bacterias inducen competencia en tales condiciones de estrés, apoya la tercera hipótesis, que la transformación existe para permitir la reparación del ADN. En pruebas experimentales, las células bacterianas expuestas a agentes que dañan su ADN y luego se transforman, sobrevivieron mejor que las células expuestas a daños en el ADN que no sufrieron transformación. Además, la competencia para someterse a la transformación a menudo es inducible por agentes que dañan el ADN conocidos. Por tanto, una fuerte ventaja selectiva a corto plazo para la competencia natural y la transformación sería su capacidad para promover la reparación del ADN recombinacional homólogo en condiciones de estrés. Estas condiciones de estrés pueden producirse durante la infección bacteriana de un huésped susceptible. Entre diferentes cepas de S. pneumoniae altamente transformables, la capacidad de colonización nasal y la virulencia (infectividad pulmonar) dependen de un sistema de competencia intacto.
Se hizo un contraargumento basado en el descubrimiento que el daño monocatenario y bicatenario del ADN cromosómico no inducía ni mejoraba la competencia o transformación en B. subtilis o H. influenzae, lo que sugiere que la selección para la reparación ha influido poco o nada en la evolución de la competencia en estas especies.
Sin embargo, la evidencia más reciente indica que la competencia para la transformación es inducida específicamente por condiciones que dañan el ADN. Por ejemplo, los agentes que dañan el ADN mitomicina C (un agente de entrecruzamiento del ADN) y fluoroquinolona (un inhibidor de la topoisomerasa que causa roturas de doble hebra) inducen la transformación en Streptococcus pneumoniae. Además, en S. pneumoniae la transformación protege contra el efecto bactericida de la mitomicina C. La inducción de la competencia protege aún más contra los antibióticos kanomicina y estreptomicina. Aunque estos antibióticos aminoglucósidos se consideraban anteriormente como no dañinos para el ADN, se mostró que una parte sustancial de su actividad bactericida es el resultado de la liberación del radical hidroxilo y la inducción de daños en el ADN, incluidas las roturas de doble cadena.
La ciprofloxacina, que interactúa con la ADN girasa y causa la producción de roturas de doble hebra, induce la expresión de genes de competencia en Helicobacter pylori, lo que conduce a una mayor transformación. Estudios de Legionella pneumophila, probaron 64 moléculas tóxicas para determinar cuáles inducen competencia. Solo seis de estas moléculas, todos agentes que dañan el ADN, indujeron fuertemente la competencia. Estas moléculas fueron norfloxacina, ofloxacina y ácido nalidíxico (inhibidores de la ADN girasa que producen roturas de doble cadena), mitomicina C (que produce enlaces cruzados entre cadenas), biciclomicina (provoca roturas de cadena simple y doble) e hidroxiurea (provoca la oxidación de las bases del ADN). También se ha demostrado que la irradiación UV induce competencia en L. pneumophila y sugirió además que la competencia para la transformación evolucionó como respuesta al daño del ADN.
Transferencia horizontal de genes
Ocasionalmente se puede conferir una ventaja a largo plazo por instancias ocasionales de transferencia horizontal de genes, también llamada transferencia lateral de genes (que puede resultar de una recombinación no homóloga después de que se induce la competencia), que podrían proporcionar resistencia a los antibióticos u otras ventajas.
Independientemente de la naturaleza de la selección por competencia, la naturaleza compuesta de los genomas bacterianos proporciona abundantes pruebas de que la transferencia horizontal de genes causada por la competencia contribuye a la diversidad genética que hace posible la evolución.
Véase también
| Control de autoridades |
|
|---|
-
 Datos: Q1780863
Datos: Q1780863
-
 Multimedia: Natural competence / Q1780863
Multimedia: Natural competence / Q1780863