
| Esponjas | ||
|---|---|---|
| Rango temporal: 760–0Ma Tónico – Reciente | ||

| ||
| Taxonomía | ||
| Reino: | Animalia | |
| Subreino: | Parazoa | |
| Filo: |
Porifera Grant, 1836 |
|
| Clases | ||
| Sinonimia | ||
| ||

Los poríferos (Porifera), también conocidos como esponjas o esponjas de mar, son un filo de animales acuáticos que se encuentran enclavados dentro del subreino Parazoa. Son filtradores gracias a un desarrollado sistema acuífero de poros, canales y cámaras que generan corrientes de agua provocados por el movimiento de unas células flageladas: los coanocitos. Existen unas nueve mil especies de esponjas en el mundo, de las cuales solo unas ciento cincuenta viven en agua dulce. Se conocen fósiles de esponjas (una hexactinélida) desde el Período Ediacárico (Neoproterozoico o Precámbrico superior). Se consideraron plantas debido a su inmovilidad hasta que en 1765 se descubrió la existencia de corrientes internas de agua y fueron reconocidos como animales. Su digestión es intracelular. Las esponjas son el grupo hermano de todos los demás animales. Las esponjas fueron las primeras formas en ramificarse del árbol evolutivo desde el ancestro común de todos los animales.
Historia
Aristóteles (siglo IV a. C.), quien estuvo en los orígenes de la taxonomía científica de los animales, los dividió en su obra "Las partes de los animales" (en griego antiguo, Περὶ ζώων μορίων), en dos grandes grupos: "animales con sangre" y "animales sin sangre", los cuales a su vez clasificó en unidades más pequeñas.
Más tarde, en los siglos IV-V d. C., los neoplatonistas Dexipo y Armonio de Hermia comenzaron a llamar a tales organismos "zoofitos"(en griego antiguo, ζωόφυτα), clasificándolos como formas intermedias entre plantas y animales. En la Europa medieval, este término casi no se usó, pero nuevamente entró en uso en el Renacimiento, ya que fue utilizado en sus clasificaciones por zoólogos como Edward Wotton, Guillaume Rondelet, Conrad Gesner y Ulisse Aldrovandi.
Las esponjas aparecían invariablemente en los "zoofitos", aunque el volumen del grupo cambiaba; así, en "De differentiis animalium" (1552) de Wotto, las estrellas de mar, los escifozoos y los ctenophores fueron incuidos a los zoofitos. En el libro de Carlos Linneo "Systema Naturae", la clase Vermes incluía el orden Zoofita con un volumen aún mayor que el de Wotton: Linneo también incluía urnas marinas y algunos moluscos y gusanos en el número de zoofitos. Al mismo tiempo, en la 10.ª edición del libro (1758), Linneo definió Zoophyta como "plantas con flores que viven la vida animal", y en la decimosegunda edición (1766-1768) como "animales complejos que florecen como las flores de las plantas".
La naturaleza animal de las esponjas fue corroborada por primera vez por John Ellis, quien describió su capacidad para crear corrientes de agua y cambiar el diámetro de los Osculum y esbozó sus observaciones en 1765 en una carta a Daniel Solander (en 1766 la carta fue publicada en Philosophical Transactions of the Royal Society). En 1752, antes del descubrimiento de Ellis, el naturalista francés Jean-André Peysonnel planteó la hipótesis de que las esponjas no son organismos vivos, sino estructuras erigidas por gusanos de mar.

La posición sistemática de las esponjas entre los "zoofitos" fue aclarada gradualmente. En Système des animaux sans vertèbres (1801) y Philosophie zoologique (1809) de Jean-Baptiste Lamarck, las esponjas aparecen como parte del orden Polyps con Polypnyak, junto con briozoos, conchas y una serie de grupos de cavidades intestinales. Georges Cuvier en su obra "Le régne animal distribué d'apres son organisation" (1817) incluyó esponjas junto con anthozoos en la clase Polypae de la sección Radiata (esta última corresponde aproximadamente a Zoophyta en el sentido de Wotton o Linneo, aunque Cuvier ya no consideraba Radiata como una transición entre plantas y organismos animales).

La transición a un estudio en profundidad de la biología de las esponjas surgió en los estudios de Robert Edmund Grant, en los que propuso la primera interpretación precisa de la estructura y la fisiología de estos animales. Ya en su primer artículo (1825), Grant describió larvas y huevos en esponjas del género Spongilla, y llevó a cabo un estudio microscópico exhaustivo de la estructura de los poros de las esponjas. En 1836, introdujo el nombre Porophora para las esponjas, que fue substituido por sí mismo en 1841 por Poriphera, y por Rymer Jonesen 1847 por Porifera (todos los nombres significan "llevando poros").
Durante la mayor parte del siglo XIX, las esponjas se asociaron generalmente con los centéreos y, a menudo, simplemente se incluían en estos últimas (aunque naturalistas como Henri Dutrochet, Paul Gervais y John Hogg clasificaron las esponjas como plantas). Sin embargo, ya en 1816, Henri de Blainville propuso la idea de una estrecha conexión entre las esponjas y los protozoos. En 1860, Hogg, y en 1866 Ernst Haeckel, propusieron asignar un reino separado (según Hogg: Protoctista, según Haeckel: Protista), el cual incluía esponjas, animales unicelulares y parte de las plantas unicelulares (Thomas Huxley, quien consideraba que las esponjas eran colonias de organismos unicelulares, en 1864 simplemente las incluyó en los protozoos) Este punto de vista parecía haber sido confirmado en el descubrimiento por Henry James-Clark en 1867 de flagelos de collar (coanoflagelados), notablemente similares a las células especiales de todas las esponjas: los coanocitos.

Sin embargo, el último tercio del siglo XIX se convirtió, en palabras de A. V. Ereskovsky, en la "edad de oro" de la embriología de las esponjas (y la primera etapa de los estudios sistemáticos de su desarrollo). Desde 1874 hasta 1879, la investigación de II. Mechnikov, Franz Schulze y Oscar Schmidt, dedicada al estudio de la estructura y el desarrollo de las esponjas, demostró irrefutablemente su pertenencia a animales multicelulares. Al mismo tiempo, se descubrió una gran originalidad de este grupo de animales. En particular, Schulze (1878) e Yves Delage (1892) describieron el llamado fenómeno de "perversión de las hojas germinales" en el desarrollo de esponjas, distinguiendo claramente a Porifera de todos los demás Metazoos (incluidos los celentéreos); sin embargo, a finales del siglo XX y principios del siglo XXI, la terminología cambió: comenzó a prevalecer la opinión según la cual las hojas germinales de las esponjas en el curso de la embriogénesis no se forman en absoluto, y por lo tanto no tiene sentido hablar de su "perversión". Por lo tanto, William Sollas (1884) contrastó las esponjas como Parazoa, separándolas de todos los demás Metazoos (los cuales pronto serían llamados Eumetazoa). En el sistema Haeckel tardío (1894-1895), las esponjas ya fueron eliminadas de Protista y consideradas en el reino animal como un tipo independiente de Spongiae, y en el sistema de Ray Lankester (1900-1909), se atribuyen claramente a Metazoa y aparecen como un tipo de Porifera (el único en la sección de Parazoa). Este último punto de vista fue absolutamente dominante a lo largo de casi todo el siglo XX, aunque el rango de Parazoa varía entre los diferentes autores: como "sección", "superdivisión" y luego "subreino".
En las décadas de 1900 y 1960 (la segunda etapa de investigación sobre el desarrollo de esponjas, según A. V. Ereskovsky), el interés en estudiar el desarrollo de las esponjas disminuyó, aunque aparecieron importantes trabajos de Henry Wilson y Claude Levy. Alrededor de 1960, comienza la tercera etapa, que se caracteriza por el predominio de estudios ultraestructurales utilizando microscopía electrónica. A finales del siglo XX, comenzó el estudio de las características citogenéticas y moleculares-biológicas de las esponjas.
Características generales
Una de las características más sorprendentes de las esponjas es que la mayoría de las células que componen su cuerpo son totipotentes, es decir, pueden transformarse en cualquiera de los otros tipos celulares según las necesidades del animal. Por tanto, se considera que las esponjas tienen una organización celular, a diferencia del resto de metazoos cuya organización es tisular (con tejidos). Carecen de verdaderas capas embrionarias.
La forma corporal generalizada de estos animales es la de un "saco" con una abertura grande en la parte superior, el ósculo, que es por donde sale el agua, y muchos poros más o menos pequeños en las paredes, que es por donde entra el agua. La filtración del alimento se produce en la cámara interna del animal, y es llevada a cabo por un tipo celular especializado y único de los poríferos, los coanocitos. Estas células tienen una gran similitud con los protozoos coanoflagelados, por lo que hoy hay pocas dudas de que están relacionados filogenéticamente. Las esponjas, los metazoos más primitivos, tuvieron probablemente un antecesor común con los coanoflagelados coloniales, similares tal vez a los actuales Proterospongia o Sphaeroeca que son simples agregados de animales unicelulares.
Las esponjas son prácticamente incapaces de desplazarse; muchas carecen de simetría corporal y por tanto no tienen una forma definida; las hay que crecen indefinidamente hasta que topan con otra esponja en crecimiento u otro obstáculo, otras que se incrustan en rocas, horadándolas, etc. Una especie determinada puede adoptar diferentes aspectos según las condiciones ambientales, como la naturaleza e inclinación del sustrato, disponibilidad de espacio, corrientes de agua, etc.
No obstante, según estudios han demostrado que algunas esponjas pueden desplazarse sobre el sustrato de un sitio a otro, pero dada su extrema lentitud (unos 4 mm al día) el fenómeno había pasado inadvertido.
La excreción, básicamente amoníaco, y el intercambio gaseoso se producen por difusión simple, sobre todo a través del coanodermo.
Las esponjas carecen de boca y de aparato digestivo y a diferencia del resto de metazoos dependen de la digestión intracelular, con lo que la fagocitosis y pinocitosis son los mecanismos utilizados para la ingestión de alimento. Tampoco tienen células nerviosas, son los únicos animales que carecen de sistema nervioso.

Anatomía
Pinacodermo

Externamente, las esponjas están cubiertas por una capa de células pseudoepiteliales anchas y largas denominadas pinacocitos; no se trata de auténtico epitelio, ya que carece de lámina basal. El conjunto de pinacocitos forman el pinacodermo o ectosoma que es análogo a la epidermis de los eumetazoos. El pinacodermo está atravesado por numerosos poros dermales tapizados cada uno por una célula arrollada llamada porocito; el agua es atraída hacia ellos y penetra en el interior. En algunas especies hay una cutícula, una capa de colágeno consistente que recubre y eventualmente sustituye al pinacodermo.
Coanodermo
La superficie interna de una esponja está tapizada por células flageladas que en su conjunto forman el coanodermo. La cavidad central principal es el espongocele o atrio. Dichas células flageladas, denominadas coanocitos, que son virtualmente idénticas a los protozoos coanoflagelados, producen la corriente de agua y son importantes en la alimentación. El coanodermo puede tener el grosor de una célula (organización asconoide), puede replegarse (organización siconoide) o puede subdividirse para formar racimos de cámaras de coanocitos independientes (organización leuconoide).
Mesohilo
Entre estas dos capas se encuentra un área organizada laxamente, el mesohilo, en la cual se pueden encontrar fibras de soporte, espículas del esqueleto y una variedad de células ameboides de gran importancia en la digestión, secreción del esqueleto, producción de gametos y el transporte de nutrientes y desechos. Los diferentes elementos del mesohilo están inmersos en un mesoglea coloidal.

Esqueleto
En el mesohilo existen numerosas fibras elásticas de colágeno (parte proteica del esqueleto) y espículas silíceas (dióxido de silicio hidratado) o calcáreas (carbonato cálcico), dependiendo de la clase a la que pertenezca, que son la parte mineral del esqueleto y lo que le da dureza. La rigidez de esta pared corporal variará según haya más parte proteica (más flexible) o más parte mineral (más dura y rígida).
Las fibras de colágeno son de dos naturalezas básicas; fibras delgadas y dispersas, y fibras de espongina, más gruesa, que forman un retículo o entramado; se hallan entrelazadas unas con otras y con las espículas, y pueden englobar granos de arena y restos de espículas del sedimento.
Las espículas calcáreas son de morfología poco variada, pero las silíceas presentan formas y tamaños diversos, distinguiéndose las megascleras (> 100 μm) de las microscleras (< 100 μm).
Con frecuencia, espículas y fibras no están dispuestas al azar sino que forman estructuras ordenadas variadas.
Tipos celulares
Dado que las esponjas carecen de auténticos tejidos y órganos, las diferentes funciones del animal son llevadas a cabo por diversos tipos celulares más o menos independientes e intercambiables.
- Pinacocitos. Los pinacocitos típicos forman el recubrimiento externo de la mayoría de las esponjas; tiene función protectora y también fagocitan.
- Basopinacocitos. Son pinacocitos especiales situados en la base de la esponja que segregan fibras que anclan la esponja al sustrato.
- Porocitos. Son células cilíndricas del pinacodermo con un canal central regulable que deja pasar más o menos volumen de agua hacia el interior. Son exclusivos de esponjas calcáreas.
- Coanocitos. Los coanocitos son las células más características de las esponjas. Están provistas de un largo flagelo central rodeado de una corona o collar, simple o doble, de microvellosidades conectadas entre sí por filamentos mucosos que forman un retículo. Los flagelos, dirigidos hacia las cámaras vibratiles, provocan corrientes de agua gracias a movimientos que, aunque no son coordinados en el tiempo, sí lo son en la dirección. El agua cargada de partículas (bacterias, fitoplancton y materia orgánica en suspensión) atraviesa las microvellosidades, donde queda atrapado el alimento que será después fagocitado.
- Colenocitos y Lofocitos. Células del mesohilo que secretan fibras de colágeno dispersas que forman un entramado de sostén en la mesohilo.
- Espongiocitos. Células del mesohilo que secretan fibras gruesas de colágeno conocidas como fibras espongina, que son el principal soporte del cuerpo de muchas esponjas.
- Esclerocitos. Se encargan de la producción de espículas, tanto calcáreas como silíceas, y se desintegran cuando se completa la secreción de la espícula.
- Miocitos. Células contráctiles fusiformes situadas en el mesohilo, que se disponen alrededor del ósculo y de los canales principales. Su citoplasma es rico en microfilamentos y microtúbulos. Su respuesta es lenta y no condicionada a estímulos eléctricos, ya que en las esponjas no hay células nerviosas.
- Arqueocitos o Amebocitos. Células ameboides del mesohilo capaces de transformarse en cualquier otro tipo celular. También tienen gran importancia en los procesos de digestión, aceptando partículas fagocitadas por los coanocitos, y son el sistema de transporte y excreción de las esponjas. Dada su totipotencia, son claves en la reproducción asexual.
- Células esferulosas. Tienen función excretora; acumulan gránulos refringentes y los liberan a la corriente exhalante.
Niveles de organización
Las esponjas presentan tres niveles de organización, cada uno de los cuales aumenta considerablemente la superficie del coanodermo con el consiguiente incremento de la eficacia en la filtración; de más simple a más complejo:
- Asconoide. Esponjas tubulares, con simetría radiada, pequeñas (< 10 cm), con una cavidad central denominada espongiocele o atrio. El movimiento de los flagelos de los coanocitos fuerzan la entrada de agua en el espongiocele a través de poros que atraviesan la pared corporal. Los coanocitos, que tapizan el espongiocele, capturan las partículas suspendidas en el agua.
- Siconoide. Tienen también simetría radiada. La pared del cuerpo es más gruesa y más compleja que las asconoides; el coanodermo recubre también la cavidad atrial. Presentan canales radiales (o cámaras flageladas), unas cámaras tapizadas de coanocitos que se abren al espongiocele a través de un poro llamado apopilo. El agua entra en los canales inhalantes a través un gran número de poros dermales y después pasa a los canales radiales por aberturas minúsculas llamadas los prosopilos. Allí el alimento es ingerido por los coanocitos. Durante su desarrollo, las esponjas siconoides pasan a través de una etapa asconoide llamado olinto.
Solo unas cuantas especies de esponjas calcáreas tienen organización ascon o sicon.
- Leuconoide. La mayoría de las demosponjas tienen organización leucon, que es la que alcanza mayor complejidad. Las esponjas leuconoides carecen de simetría radiada, tienen la cavidad atrial reducida y poseen numerosas cámaras vibrátiles, cámaras globulares tapizadas de coanocitos independientes unas de otras y hundidas en el mesohilo y comunicadas entre sí, con el exterior y con el ósculo por una multitud de canales inhalantes y exhalantes. Los apopilos desembocan en canales exhalantes; los diversos canales exhalantes se reúnen para expulsar el agua a través de varios ósculos. Una esponja leuconoide grande puede tener varios ósculos, lo que puede interpretarse como una colonia de individuos o un solo individuo complejo.

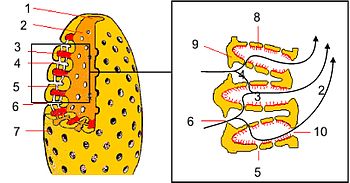

Reproducción y desarrollo
Todas las esponjas se reproducen de manera sexual, pero son muy comunes diversos tipos de reproducción asexual.
Reproducción asexual
Dada la potencialidad total de sus células, todas las esponjas pueden reproducirse asexualmente a partir de fragmentos. Muchas esponjas producen yemas, pequeñas protuberancias que acaban desprendiéndose, que en algunos casos contienen alimentos almacenados. Las especies de agua dulce (Spongillidae) producen gémulas complejas, pequeñas esferas bien organizadas con arqueocitos y varias capas protectoras, entre ellas una gruesa de colágeno sostenida por espículas de tipo anfidisco; son muy resistentes ante las inclemencias ambientales, como la desecación y la congelación (soportan -10 °C). Algunas especies marinas producen gémulas más simples, llamadas soritos.
Reproducción sexual
Las esponjas carecen de gónadas, y los gametos y los embriones se hallan en mesohilo. La mayoría son hermafroditas, pero existe una gran variabilidad, llegando al extremo que en una misma especie conviven individuos hermafroditas con individuos dioicos. En cualquier caso, la fecundación es casi siempre cruzada.
Los espermatozoides se forman a partir de coanocitos, cuando todos los de una cámara sufren la espermatogénesis y originan un quiste espermático. Los óvulos a partir de coanocitos o de arqueocitos y se rodean de una capa de células alimentarias o trofocitos. Los espermatozoides y los óvulos son expulsados al exterior a través del sistema acuífero; en este caso la fecundación se produce en el agua y da lugar a larvas planctónicas. En algunas especies, los espermatozoides penetran en el sistema acuífero de otros individuos donde son fagocitados por los coanocitos; luego, estos coanocitos se desprenden, se transforman en células ameboides (forocitos) que llevan el espermatozoide hasta un óvulo; tras la fecundación, las larvas son liberadas a través del sistema acuífero.
Existen cuatro tipos básicos de larvas en las esponjas:
- Parenquímula. Es una larva maciza, con una capa de células monoflageladas en el exterior y una masa de células parecidas a los arqueocitos en el interior inmersas en una matriz.
- Celoblástula. Es una larva hueca compuesta por una capa de células monoflageladas que rodean una cavidad interior.
- Estomoblástula. Es un tipo especial de celoblástula, propio de esponjas que incuban los embriones en su mesohilo. Es también hueca, pero posee algunas células más grandes (macrómeros) que dejan una abertura que comunica con la cavidad interior. Sufre un sorprendente proceso de inversión en que las células flageladas que un principio eran internas, acaban siendo externas.
- Anfiblástula. Es el resultado del proceso de inversión de una estomoblástula. Consta de un hemisferio formado por células grandes y no flageladas (macrómeros) y otro con células pequeñas y monoflageladas (micrómeros). La anfiblástula es liberada y acaba fijándose al sustrato por los micrómeros; éstos se invaginan formando una cámara de células flageladas que serán el futuro coanodermo; los macrómeros forman el pinacodermo; luego se abre un ósculo originando una pequeña esponja leuconoide denominada olinto.
Agregación celular
Las esponjas poseen una propiedad única y extraordinaria: cuando sus células son separadas por medios mecánicos (por ejemplo, al tamizarlas), inmediatamente vuelven a reunirse y a formar, en pocas semanas, un individuo completo y funcional; es más, si se desmenuzan dos esponjas de diferente especie, las células se separan y reagrupan volviendo a reconstruir los individuos separados.
Ecología
Por su estructura corporal (sistema acuífero de filtración), las esponjas siempre habitan el medio acuático, ya sea dulce o marino, y se fijan a un sustrato sólido, aunque algunas especies puede fijarse en sustratos blandos como arena o lodo. La mayoría de las esponjas son esciófilas (prefieren la penumbra). Su principal fuente de alimento son partículas orgánicas submicroscópicas en suspensión, muy abundantes en el mar, aunque también ingieren bacterias, dinoflagelados y otro plancton de pequeñas dimensiones. Su capacidad filtradora es destacable; una esponja leuconoide de 10 cm de altura y 1 cm de diámetro contiene 2 250 000 cámaras flageladas y filtra 22,5 litros de agua al día.
A pesar de su simplicidad, las esponjas tienen un gran éxito ecológico; son los animales dominantes en muchos hábitats bentónicos marinos y toleran bien la contaminación por hidrocarburos, metales pesados y detergentes, acumulando dichos contaminantes en grandes concentraciones sin daño aparente.
Algunas esponjas poseen simbiontes fotosintéticos (cianobacterias, zooxantelas, diatomeas, zooclorelas) o no (bacterias). Periódicamente expulsan los simbiontes y células somáticas, y secretan substancias mucosas con regularidad. En ciertas esponjas los simbiontes llegan a representar el 38% de su volumen corporal.
Son pocos los animales que se alimentan de esponjas, debido a su esqueleto de espículas y a su toxicidad. Algunos moluscos opistobranquios, equinodermos y peces. Con frecuencia se trata de especies muy específicas que son exclusivamente espongiófagas y depredan sobre una especie concreta de esponja.
Las esponjas poseen una variedad sorprendente de toxinas y antibióticos que usan para evitar la depredación y en la competencia por el sustrato. Algunas de estos compuestos se han revelado de utilidad farmacológica, con propiedades antiinflamatorias, cardiovasculares, gastrointestinales, antivíricas, antitumorales, etc., y están siendo investigadas intensamente. Entre estos compuestos se incluyen arabinósidos, terpenoides, halicondrinas, etc.
Muchos invertebrados y diversos peces utilizan las esponjas, por su estructura porosa, como lugar de residencia o refugio. Algunos gasterópodos y bivalvos tienen esponjas incrustantes sobre sus conchas y muchos cangrejos recolectan esponjas que colocan sobre sus caparazones. Se trata de casos de mutualismo, en que dichos animales consiguen camuflaje y las esponjas un método de desplazamiento.
Sistemática
El filo Porifera se divide en tres clases:
- Clase Calcarea (esponjas calcáreas). Espículas de 1, 3 o 4 radios, de carbonato cálcico cristalizado en forma de calcita. Las hay de los tres tipos de organización. En general viven en aguas costeras poco profundas.
- Clase Hexactinellida (esponjas vítreas). Espículas silíceas (dióxido de silicio hidratado) de tres o seis radios. En general viven a mayor profundidad, entre los 450 y los 900 m.
- Clase Demospongiae (demosponjas). Espículas silíceas (dióxido de silicio hidratado) monoaxonas o tetrasxonas, que pueden sustituirse por una malla de fibras de espongina. Todas tienen organización leuconoide. Viven a cualquier profundidad.
- Archaeocyatha†. Son un grupo extinto de posición incierta relacionado con las esponjas; tuvieron una corta existencia, de unos 50 millones de años, durante el Cámbrico.
La clase Sclerospongiae fue abandonada en los años 90. La constituían esponjas que producen una matriz calcárea sólida, similar a una roca, por lo que son conocidas como esponjas coralinas. Las 15 especies conocidas fueron incluidas entre las clases Calcarea y Demospongiae.
Filogenia
Monofilia
Las esponjas han sido consideradas históricamente como un grupo monofilético por su morfología y características comunes. La era del estudio molecular desafió esta visión al proponer que Porifera fuese parafilético, insinuando que los animales ancestrales fueron esponjas; sin embargo, el estudio filogenómico más profundo reconsidera la monofilia de las esponjas. Actualmente se considera que las esponjas tendrían las siguientes relaciones:
| Porifera |
|
||||||||||||||||||
Parafilia
Estudios recientes sobre la histología de las esponjas hexactinélidas han revelado que este grupo tiene peculiaridades importantes. Basándose en esto, se ha propuesto que el filo poríferos sea dividido en dos subfilos, Symplasma (hexactinélidas) y Cellularia (calcáreas + demosponjas); incluso se ha puesto en duda que los poríferos sean un grupo monofilético, es decir, que tengan todos un antepasado común y exclusivo de las esponjas (Zrzavý et al.). Los simplasmas comprenden únicamente la clase hexactinélidas; son poríferos de organización sencilla, sincitiales, aunque presentan tipos celulares como arqueocitos y células esferulosas; el pinacodermo no posee pinacocitos diferenciados y no muestran contractibilidad ya que carece de miocitos. Los celulares reúnen las clases calcáreas y demosponjas, las cuales presentan una organización celular definida, con pinacocitos y coanocitos individualizados, y diversos tipos de células ameboides en el mesohilo.
Por otro lado, estudios genéticos que sostienen que las esponjas forman un grupo parafilético dieron el siguiente resultado:
| Animalia |
|
|||||||||||||||||||||||||||||||
Porifera como el filo no basal del reino Animalia
Otros análisis basado en la secuencia de genes y aminoácidos afirman que Ctenophora sería el primer grupo en separarse del reino Animalia en contraposición con la visión tradicional de que Porifera es el grupo más basal. Este estudio sostiene que Porifera es un grupo monofilético que divergió como grupo hermano del clado ParaHoxozoa. También se ha demostrado que los poriferos comparten con estos últimos ciertas duplicaciones de genes antiguos y genes reguladores del metabolismo que están ausentes en los cténoforos. No obstante los ctenóforos son un grupo importante para entender la evolución de los animales. Si los ctenóforos realmente son el grupo hermano de los demás animales dificultaría probar como se originaron los tejidos en los animales ya que los esponjas no poseen esta característica. Sin embargo se ha demostrado que las razones por la que los ctenóforos aparecen como el grupo más basal de los animales en análisis es porque tienen una desviación en la secuencia de nucleótidos y aminoácidos lo que produce su mala colocación también llamado atracción de ramas largas. Estudios que intentan evitar el error sistemático han sugerido que las esponjas son el grupo basal de los animales.
| Apoikozoa |
|
||||||||||||||||||||||||||||||
Una característica diagnóstica de los poríferos es la presencia de espículas. Por ello, ciertos grupos extintos con organización de esponja actual fueron colocados fuera del filo Porifera. En particular, son problemáticos grupos con esqueleto calcáreo sólido como arqueociatos, caetétidos, esfinctozoos, estromatoporoides y receptacúlidos. El descubrimiento de unas quince especies vivientes con esqueleto calcáreo sólido, ha ayudado mucho a comprender la filogenia de los poríferos. Estas especies poseen formas diversas y deberían clasificarse con los caetétidos, esfinctozoos y estromatoporoides si se hubieran hallado como fósiles. No obstante, con el estudio del material vivo, las características histológicas, citológicas y larvarias muestran claramente que esas quince especies pueden situarse unas entre las esponjas calcáreas y otras entre las demosponjas. Por tanto, las formas fósiles correspondientes, muy abundantes, también encajarían en una de estas dos clases.
Está ampliamente aceptado entre los especialistas que las calcáreas y las demosponjas están más estrechamente relacionadas entre sí que con las hexactinélidas. Con el descubrimiento de las quince formas vivientes antes mencionadas, fue creada una cuarta clase, las esclerosponjas. No obstante, se trata de un grupo polifilético que debería ser abandonado, según Chombard, et al. Los arqueociatos representan un caso especial; no existen representantes vivos, aunque su organización puede referirse a la de las actuales esponjas. El análisis filogenético realizado por Reitner & Mehl los coloca como grupo hermano de las demosponjas. Por tanto, los arqueociatos perderían su categoría de filo y pasarían a ser una clase dentro de las esponjas.
Distribución

Las esponjas son comunes en todo el mundo. La mayoría de las especies habitan en aguas oceánicas desde regiones polares hasta regiones tropicales. Aun así, las esponjas alcanzan la mayor diversidad de especies en las zonas tropicales y subtropicales del Océano Mundial. 219 especies (según otras fuentes, cerca de 150) viven en cuerpos de agua dulce.
La mayoría de las especies viven en aguas tranquilas y limpias, ya que las partículas de limo y arena suspendidas en el agua o levantadas del fondo por las corrientes pueden obstruir los poros en los cuerpos de las esponjas, obstaculizando así los procesos de respiración y nutrición. Dado que las esponjas llevan un estilo de vida sedentario adjunto, necesitan un sustrato sólido para su desarrollo y crecimiento. En este sentido, las acumulaciones de esponjas ocurren en lugares donde hay materiales pedregosos en el fondo: piedras, cantos rodados, guijarros, etc. Sin embargo, algunas especies pueden fijarse en sedimentos sedimentarios blandos del fondo con la ayuda de una base en forma de raíz de su cuerpo.
La mayoría de las esponjas viven a profundidades poco profundas, hasta 100-500 m. Con el aumento de la profundidad, el número de especies de esponjas disminuye. A profundidades superiores a 1000-1500 m, las esponjas suelen ser bastante raras, de modo que el número de especies de esponjas en aguas profundas sea pequeño.
En aguas templadas, las esponjas son más numerosas, pero menos diversas que en los trópicos. Esto se debe probablemente al hecho de que en los trópicos hay muchos organismos que se alimentan de esponjas. Las esponjas vítreas son más numerosas en aguas polares, así como a grandes profundidades de mares templados y tropicales, ya que su estructura corporal porosa les permite extraer partículas de alimentos de estas aguas pobres en alimentos a un costo mínimo. Las esponjas comunes y las esponjas calcáreas son numerosas y variadas en aguas no polares más tranquilas.
Las esponjas que viven en zonas de marea están bien adaptadas a una corta estancia al aire libre, cuando la marea baja sobresalen del agua. Sus bocas y poros están cerrados, lo que evita la pérdida excesiva de humedad y el secado.
Antigüedad
Las esponjas vienen a ser el grupo más antiguo de animales vivientes. De acuerdo con el registro fósil, aparecen aproximadamente hace unos 760 millones de años, en el Tónico antes de la fauna ediacárica. Este fósil temprano se conoce como Otavia.
Uso
Por delfines
Un informe en 1997 describió el uso de esponjas como herramienta por los delfines en la bahía Shark en Australia Occidental. Un delfín uniría una esponja marina a su rostro, el cual presumiblemente utilizaría para protegerse cuando busca alimento en el fondo marino arenoso. Este comportamiento sólo se ha observado en esta bahía, y es mostrado casi exclusivamente por las hembras. Un estudio en 2005 concluyó que las madres enseñan el comportamiento a sus hijas, y que todas las esponjas están estrechamente relacionados, lo que sugiere que es una innovación bastante reciente.
Por humanos
Comercio y explotación

Los primeros habitantes del Mediterráneo ya utilizaban la conocida esponja de baño; su uso fue descubierto probablemente por los egipcios. Aristóteles conocía las esponjas y describió su gran capacidad de regeneración. Los soldados romanos usaban esponjas en vez de copas de metal para beber agua durante las campañas militares, y la pesca de esponjas era una de las pruebas de los antiguos juegos olímpicos.
En el Atlántico Norte se han usado tradicionalmente las esponjas arrojadas a las playas por el mar como fertilizante para los campos de cultivo. No obstante, el interés económico radica en las esponjas de baño, sobre todo los géneros Spongia e Hippospongia, cuyo esqueleto es exclusivamente córneo y flexible. El comercio de esponjas se ha centrado durante años en el Mediterráneo Oriental, las costas del Atlántico americano, del golfo de México y el Caribe hacia el norte, y Japón. En Florida se hallaba la industria manufacturera más importante del mundo. A mediados del siglo XX, la pesca abusiva y diversas epidemias redujeron drásticamente el volumen de esponjas comercializado. La decadencia de este comercio se vio acentuada con la aparición de esponjas sintéticas.
Algunas demosponjas son perjudiciales para el hombre ya que perforan las conchas de los moluscos, produciendo daños en los criaderos de bivalvos (mejillones, ostras, etc.)
Medicina
Las esponjas tienen potencial medicinal debido a la presencia en las propias esponjas o en sus simbiontes microbianos de sustancias químicas que pueden ser utilizadas para controlar virus, bacterias, tumores y hongos.
Fuentes
- Grant, Robert Edmund (1836). «Animal Kingdom». En Todd, R. B., ed. The Cyclopaedia of Anatomy and Physiology. Volume 1. Londres: Sherwood,Gilbert, and Piper. pp. 107-118.
Enlaces externos
-
 Wikimedia Commons alberga una galería multimedia sobre Porifera.
Wikimedia Commons alberga una galería multimedia sobre Porifera. -
 Wikispecies tiene un artículo sobre Porifera.
Wikispecies tiene un artículo sobre Porifera.
| Control de autoridades |
|
|---|